Assessing bat-vehicle collision risks using acoustic 3D tracking
Recommended by Gloriana Chaverri based on reviews by Mark Brigham and ? based on reviews by Mark Brigham and ?
The loss of biodiversity is an issue of great concern, especially if the extinction of species or the loss of a large number of individuals within populations results in a loss of critical ecosystem services. We know that the most important threat to most species is habitat loss and degradation (Keil et al., 2015; Pimm et al., 2014); the latter can be caused by multiple anthropogenic activities, including pollution, introduction of invasive species and fragmentation (Brook et al., 2008; Scanes, 2018). Roads are a major cause of habitat fragmentation, isolating previously connected populations and being a direct source of mortality for animals that attempt to cross them (Spellberg, 1998).
While most studies have focused on the effect of roads on larger mammals (Bartonička et al., 2018; Litvaitis and Tash, 2008), in recent years many researchers have grown increasingly concerned about the risk of collision between bats and vehicles (Fensome and Mathews, 2016). For example, a recent publication by Medinas et al. (2021) found 509 bat casualties along a 51-km-long transect during a period of 3 years. Their study provides extremely valuable information to asses which factors primarily drive bat mortality on roads, yet it required a substantial investment of time coupled with the difficulty of detecting bat carcasses. Other studies have used acoustic monitoring as a proxy to gauge risk of collision based on estimates of bat density along roads (reviewed in Fensome and Mathews 2016); while the results of such studies are valuable, the number of passes recorded does not necessarily equal collision risk, as many species may simply avoid crossing the roads. Understanding the risk of collisions is of vital importance for adequate planning of road construction, particularly for key sites that harbor threatened bat species or unusually large populations, especially if these are already greatly impacted by other anthropogenic activities (e.g. wind turbines; Kunz et al. 2007) or unusually deadly pathogens (e.g. white-nose syndrome; Blehert et al. 2009).
The study by Roemer et al. (2020) titled “Influence of local landscape and time of year on bat-road collision risks”, is a welcome addition to our understanding of bat collision risk as it employs a more accurate assessment of bat collision risk based on acoustic monitoring and tracking of flight paths. The goal of the study of Roemer and collaborators, which was conducted at 66 study sites in the Mediterranean region, is to provide an assessment of collision risk based on bat activity near roads. They collected a substantial amount of information for several species: more than 30,000 estimated flight trajectories for 21+ species, including Barbastella barbastellus, Myotis spp., Plecotus sp., Rhinolophus ferrumequinum, Miniopterus schreibersii, Pipistrellus spp., Nyctalus leisleri, and others. They assess risk based on estimates of 1) species abundance from acoustic monitoring, 2) direction of flight paths along roads, and 3) bat-vehicle co-occurrence.
Their findings suggest that risk is habitat, species, guild, and season-specific. Roads within forested habitats posed the largest threats for most species, particularly since most flights within these habitats occurred at the zone of collision risk. They also found that bats typically fly parallel to the road axis regardless of habitat type, which they argue supports the idea that bats may use roads as corridors. The results of their study, as expected, also show that the majority of bat passes were detected during summer or autumn, depending on species, yet they provide novel findings of an increase in risky behaviors during autumn, when the number of passes at the zone of collision risk increased significantly. Their results also suggest that mid-range echolocators, a classification that is based on call design and parameters (Frey-Ehrenbold et al., 2013), had a larger portion of flights in the zone at risk, thus potentially making them more susceptible than short and long-range echolocators to collisions with vehicles.
The methods employed by Roemer et al. (2020) could further help us determine how roads pose species and site-specific threats in a diversity of places without the need to invest a significant amount of time locating bat carcasses. Their findings are also important as they could provide valuable information for deciding where new roads should be constructed, particularly if the most vulnerable species are abundant, perhaps due to the presence of important roost sites. They also show how habitats near larger roads could increase threats, providing an important first step for recommendations regarding road construction and maintenance. As pointed out by one reviewer, one possible limitation of the study is that the results are not supported by the identification of carcasses. For example, does an increase in the number of identified flights at the zone of risk really translate into an increase in the number of collisions? Regardless of the latter, the paper’s methods and results are very valuable and provide an important step towards developing additional tools to assess bat-vehicle collision risks.
References
[1] Bartonička T, Andrášik R, Duľa M, Sedoník J, Bíl M (2018) Identification of local factors causing clustering of animal-vehicle collisions. The Journal of Wildlife Management, 82, 940–947. https://doi.org/10.1002/jwmg.21467
[2] Blehert DS, Hicks AC, Behr M, Meteyer CU, Berlowski-Zier BM, Buckles EL, Coleman JTH, Darling SR, Gargas A, Niver R, Okoniewski JC, Rudd RJ, Stone WB (2009) Bat White-Nose Syndrome: An Emerging Fungal Pathogen? Science, 323, 227–227. https://doi.org/10.1126/science.1163874
[3] Brook BW, Sodhi NS, Bradshaw CJA (2008) Synergies among extinction drivers under global change. Trends in Ecology & Evolution, 23, 453–460. https://doi.org/10.1016/j.tree.2008.03.011
[4] Fensome AG, Mathews F (2016) Roads and bats: a meta-analysis and review of the evidence on vehicle collisions and barrier effects. Mammal Review, 46, 311–323. https://doi.org/10.1111/mam.12072
[5] Frey‐Ehrenbold A, Bontadina F, Arlettaz R, Obrist MK (2013) Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matrices. Journal of Applied Ecology, 50, 252–261. https://doi.org/10.1111/1365-2664.12034
[6] Keil P, Storch D, Jetz W (2015) On the decline of biodiversity due to area loss. Nature Communications, 6, 8837. https://doi.org/10.1038/ncomms9837
[7] Kunz TH, Arnett EB, Erickson WP, Hoar AR, Johnson GD, Larkin RP, Strickland MD, Thresher RW, Tuttle MD (2007) Ecological impacts of wind energy development on bats: questions, research needs, and hypotheses. Frontiers in Ecology and the Environment, 5, 315–324. https://doi.org/10.1890/1540-9295(2007)5[315:EIOWED]2.0.CO;2
[8] Litvaitis JA, Tash JP (2008) An Approach Toward Understanding Wildlife-Vehicle Collisions. Environmental Management, 42, 688–697. https://doi.org/10.1007/s00267-008-9108-4
[9] Medinas D, Marques JT, Costa P, Santos S, Rebelo H, Barbosa AM, Mira A (2021) Spatiotemporal persistence of bat roadkill hotspots in response to dynamics of habitat suitability and activity patterns. Journal of Environmental Management, 277, 111412. https://doi.org/10.1016/j.jenvman.2020.111412
[10] Pimm SL, Jenkins CN, Abell R, Brooks TM, Gittleman JL, Joppa LN, Raven PH, Roberts CM, Sexton JO (2014) The biodiversity of species and their rates of extinction, distribution, and protection. Science, 344. https://doi.org/10.1126/science.1246752
[11] Roemer C, Coulon A, Disca T, Bas Y (2020) Influence of local landscape and time of year on bat-road collision risks. bioRxiv, 2020.07.15.204115, ver. 3 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2020.07.15.204115
[12] Scanes CG (2018) Chapter 19 - Human Activity and Habitat Loss: Destruction, Fragmentation, and Degradation. In: Animals and Human Society (eds Scanes CG, Toukhsati SR), pp. 451–482. Academic Press. https://doi.org/10.1016/B978-0-12-805247-1.00026-5
[13] Spellerberg I (1998) Ecological effects of roads and traffic: a literature review. Global Ecology & Biogeography Letters, 7, 317–333. https://doi.org/10.1046/j.1466-822x.1998.00308.x
| Influence of local landscape and time of year on bat-road collision risks | Charlotte Roemer, Aurélie Coulon, Thierry Disca, and Yves Bas | <p>Roads impact bat populations through habitat loss and collisions. High quality habitats particularly increase bat mortalities on roads, yet many questions remain concerning how local landscape features may influence bat behaviour and lead to hi... |  | Behaviour & Ethology, Biodiversity, Conservation biology, Human impact, Landscape ecology | Gloriana Chaverri | | 2020-07-20 10:56:29 | View |
Hough transform implementation to evaluate the morphological variability of the moon jellyfish (Aurelia spp.)
Céline Lacaux, Agnès Desolneux, Justine Gadreaud, Bertrand Martin-Garin and Alain Thiéry
https://doi.org/10.1101/2020.03.11.986984
A new member of the morphometrics jungle to better monitor vulnerable lagoons
Recommended by Vincent Bonhomme based on reviews by Julien Claude and 1 anonymous reviewer
In the recent years, morphometrics, the quantitative description of shape and its covariation [1] gained considerable momentum in evolutionary ecology. Using the form of organisms to describe, classify and try to understand their diversity can be traced back at least to Aristotle. More recently, two successive revolutions rejuvenated this idea [1–3]: first, a proper mathematical refoundation of the theory of shape, then a technical revolution in the apparatus able to acquire raw data. By using a feature extraction method and planning its massive use on data acquired by aerial drones, the study by Lacaux and colleagues [4] retraces this curse of events.
The radial symmetry of Aurelia spp. jelly fish, a common species complex, is affected by stress and more largely by environmental variations, such as pollution exposition. Aurelia spp. normally present four gonads so that the proportion of non-tetramerous individuals in a population has been proposed as a biomarker [5,6].
In this study, the authors implemented the Hough transform to largely automate the detection of the gonads in Aurelia spp. Such use of the Hough transform, a long-used approach to identify shapes through edge detection, is new to morphometrics. Here, the Aurelia spp. gonads are identified as ellipses from which aspect descriptors can be derived, and primarily counted and thus can be used to quantify the proportion of individuals presenting body plans disorders.
The sample sizes studied here were too low to allow finer-grained ecophysiological investigations. That being said, the proof-of-concept is convincing and this paper paths the way for an operational and innovative approach to the ecological monitoring of sensible aquatic ecosystems.
References
[1] Kendall, D. G. (1989). A survey of the statistical theory of shape. Statistical Science, 87-99. doi: https://doi.org/10.1214/ss/1177012589
[2] Rohlf, F. J., and Marcus, L. F. (1993). A revolution morphometrics. Trends in ecology & evolution, 8(4), 129-132. doi: https://doi.org/10.1016/0169-5347(93)90024-J
[3] Adams, D. C., Rohlf, F. J., and Slice, D. E. (2004). Geometric morphometrics: ten years of progress following the ‘revolution’. Italian Journal of Zoology, 71(1), 5-16. doi: https://doi.org/10.1080/11250000409356545
[4] Lacaux, C., Desolneux, A., Gadreaud, J., Martin-Garin, B. and Thiéry, A. (2020) Hough transform implementation to evaluate the morphological variability of the moon jellyfish (Aurelia spp.). bioRxiv, 2020.03.11.986984, ver. 3 peer-reviewed and recommended by Peer Community in Ecology. doi: https://doi.org/10.1101/2020.03.11.986984
[5] Gershwin, L. A. (1999). Clonal and population variation in jellyfish symmetry. Journal of the Marine Biological Association of the United Kingdom, 79(6), 993-1000. doi: https://doi.org/10.1017/S0025315499001228
[6] Gadreaud, J., Martin-Garin, B., Artells, E., Levard, C., Auffan, M., Barkate, A.-L. and Thiéry, A. (2017) The moon jellyfish as a new bioindicator: impact of silver nanoparticles on the morphogenesis. In: Mariottini GL, editor. Jellyfish: ecology, distribution patterns and human interactions. Nova Science Publishers; 2017. pp. 277–292.
| Hough transform implementation to evaluate the morphological variability of the moon jellyfish (Aurelia spp.) | Céline Lacaux, Agnès Desolneux, Justine Gadreaud, Bertrand Martin-Garin and Alain Thiéry | <p>Variations of the animal body plan morphology and morphometry can be used as prognostic tools of their habitat quality. The potential of the moon jellyfish (Aurelia spp.) as a new model organism has been poorly tested. However, as a tetramerous... | 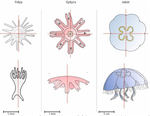 | Morphometrics | Vincent Bonhomme | | 2020-03-18 17:40:51 | View |
Once upon a time in the far south: Influence of local drivers and functional traits on plant invasion in the harsh sub-Antarctic islands
Manuele Bazzichetto, François Massol, Marta Carboni, Jonathan Lenoir, Jonas Johan Lembrechts, Rémi Joly, David Renault
https://doi.org/10.1101/2020.07.19.210880
A meaningful application of species distribution models and functional traits to understand invasion dynamics
Recommended by Joaquín Hortal based on reviews by Paula Matos and Peter Convey
Polar and subpolar regions are fragile environments, where the introduction of alien species may completely change ecosystem dynamics if the alien species become keystone species (e.g. Croll, 2005). The increasing number of human visits, together with climate change, are favouring the introduction and settling of new invaders to these regions, particularly in Antarctica (Hughes et al. 2015). Within this context, the joint use of Species Distribution Models (SDM) –to assess the areas potentially suitable for the aliens– with other measures of the potential to become successful invaders can inform on the need for devoting specific efforts to eradicate these new species before they become naturalized (e.g. Pertierra et al. 2016). Bazzichetto et al. (2020) use data from a detailed inventory, SDMs and trait data altogether to assess the drivers of invasion success of six alien plants on Possession Island, in the remote sub-Antarctic archipelago of Crozet. SDMs have inherent limitations to describe different aspects of species distributions, including the fundamental niche and, with it, the areas that could host viable populations (Hortal et al. 2012). Therefore, their utility to predict future biological invasions is limited (Jiménez-Valverde et al. 2011). However, they can be powerful tools to describe species range dynamics if they are thoughtfully used by adopting conscious decisions about the techniques and data used, and interpreting carefully the actual implications of their results. This is what Bazzichetto et al. (2020) do, using General Linear Models (GLM) –a technique well rooted in the original niche-based SDM theory (e.g. Austin 1990)– that can provide a meaningful description of the realized niche within the limits of an adequately sampled region. Further, as alien species share and are similarly affected by several steps of the invasion process (Richardson et al. 2000), these authors model the realized distribution of the six species altogether. This can be done through the recently developed joint-SDM, a group of techniques where the co-occurrence of the modelled species is explicitly taken into account during modelling (e.g. Pollock et al. 2014). Here, the addition of species traits has been identified as a key step to understand the associations of species in space (see Dormann et al. 2018). Bazzichetto et al. (2020) combine their GLM-based SDM for each species with a so-called multi-SDM approach, where they assess together the consistency in the interactions between both species and topographically-driven climate variations, and several plant traits and two key anthropic factors –accessibility from human settlements and distance to hiking paths. This work is a good example on how a theoretically meaningful SDM approach can provide useful –though perhaps not deep– insights on biological invasions for remote landscapes threatened by biotic homogenization. By combining climate and topographic variables as proxies for the spatial variations in the abiotic conditions regulating plant growth, measures of accessibility, and traits of the plant invaders, Bazzichetto et al. (2020) are able to identify the different effects that the interactions between the potential intensity of propagule dissemination by humans, and the ecological characteristics of the invaders themselves, may have on their invasion success. The innovation of modelling together species responses is important because it allows dissecting the spatial dynamics of spread of the invaders, which indeed vary according to a handful of their traits. For example, their results show that no all old residents have profited from the larger time of residence in the island, as Poa pratensis is seemingly as dependent of a higher intensity of human activity as the newcomer invaders in general are. According to Bazzichetto et al. trait-based analyses, these differences are apparently related with plant height, as smaller plants disperse more easily. Further, being perennial also provides an advantage for the persistence in areas with less human influence. This puts name, shame and fame to the known influence of plant life history on their dispersal success (Beckman et al. 2018), at least for the particular case of plant invasions in Possession Island. Of course this approach has limitations, as data on the texture, chemistry and temperature of the soil are not available, and thus were not considered in the analyses. These factors may be critical for both establishment and persistence of small plants in the harsh Antarctic environments, as Bazzichetto et al. (2020) recognize. But all in all, their results provide key insights on which traits may confer alien plants with a higher likelihood of becoming successful invaders in the fragile Antarctic and sub-Antarctic ecosystems. This opens a way for rapid assessments of invasibility, which will help identifying which species in the process of naturalizing may require active contention measures to prevent them from becoming ecological game changers and cause disastrous cascade effects that shift the dynamics of native ecosystems. **References** Austin, M. P., Nicholls, A. O., and Margules, C. R. (1990). Measurement of the realized qualitative niche: environmental niches of five Eucalyptus species. Ecological Monographs, 60(2), 161-177. doi: [https://doi.org/10.2307/1943043](https://doi.org/10.2307/1943043) Bazzichetto, M., Massol, F., Carboni, M., Lenoir, J., Lembrechts, J. J. and Joly, R. (2020) Once upon a time in the far south: Influence of local drivers and functional traits on plant invasion in the harsh sub-Antarctic islands. bioRxiv, 2020.07.19.210880, ver. 3 peer-reviewed and recommended by PCI Ecology. doi: [https://doi.org/10.1101/2020.07.19.210880](https://doi.org/10.1101/2020.07.19.210880) Beckman, N. G., Bullock, J. M., and Salguero-Gómez, R. (2018). High dispersal ability is related to fast life-history strategies. Journal of Ecology, 106(4), 1349-1362. doi: [https://doi.org/10.1111/1365-2745.12989](https://doi.org/10.1111/1365-2745.12989) Croll, D. A., Maron, J. L., Estes, J. A., Danner, E. M., and Byrd, G. V. (2005). Introduced predators transform subarctic islands from grassland to tundra. Science, 307(5717), 1959-1961. doi: [https://doi.org/10.1126/science.1108485](https://doi.org/10.1126/science.1108485) Dormann, C. F., Bobrowski, M., Dehling, D. M., Harris, D. J., Hartig, F., Lischke, H., Moretti, M. D., Pagel, J., Pinkert, S., Schleuning, M., Schmidt, S. I., Sheppard, C. S., Steinbauer, M. J., Zeuss, D., and Kraan, C. (2018). Biotic interactions in species distribution modelling: 10 questions to guide interpretation and avoid false conclusions. Global Ecology and Biogeography, 27(9), 1004-1016. doi: [https://doi.org/10.1111/geb.12759](https://doi.org/10.1111/geb.12759) Jiménez-Valverde, A., Peterson, A., Soberón, J., Overton, J., Aragón, P., and Lobo, J. (2011). Use of niche models in invasive species risk assessments. Biological Invasions, 13(12), 2785-2797. doi: [https://doi.org/10.1007/s10530-011-9963-4](https://doi.org/10.1007/s10530-011-9963-4) Hortal, J., Lobo, J. M., and Jiménez-Valverde, A. (2012). Basic questions in biogeography and the (lack of) simplicity of species distributions: Putting species distribution models in the right place. Natureza & Conservação – Brazilian Journal of Nature Conservation, 10(2), 108-118. doi: [https://doi.org/10.4322/natcon.2012.029](https://doi.org/10.4322/natcon.2012.029) Hughes, K. A., Pertierra, L. R., Molina-Montenegro, M. A., and Convey, P. (2015). Biological invasions in terrestrial Antarctica: what is the current status and can we respond? Biodiversity and Conservation, 24(5), 1031-1055. doi: [https://doi.org/10.1007/s10531-015-0896-6](https://doi.org/10.1007/s10531-015-0896-6) Pertierra, L. R., Baker, M., Howard, C., Vega, G. C., Olalla-Tarraga, M. A., and Scott, J. (2016). Assessing the invasive risk of two non-native Agrostis species on sub-Antarctic Macquarie Island. Polar Biology, 39(12), 2361-2371. doi: [https://doi.org/10.1007/s00300-016-1912-3](https://doi.org/10.1007/s00300-016-1912-3) Pollock, L. J., Tingley, R., Morris, W. K., Golding, N., O'Hara, R. B., Parris, K. M., Vesk, P. A., and McCarthy, M. A. (2014). Understanding co-occurrence by modelling species simultaneously with a Joint Species Distribution Model (JSDM). Methods in Ecology and Evolution, 5(5), 397-406. doi: [https://doi.org/10.1111/2041-210X.12180](https://doi.org/10.1111/2041-210X.12180) Richardson, D. M., Pyšek, P., Rejmánek, M., Barbour, M. G., Panetta, F. D., and West, C. J. (2000). Naturalization and invasion of alien plants: concepts and definitions. Diversity and Distributions, 6(2), 93-107. doi: [https://doi.org/10.1046/j.1472-4642.2000.00083.x](https://doi.org/10.1046/j.1472-4642.2000.00083.x)
| Once upon a time in the far south: Influence of local drivers and functional traits on plant invasion in the harsh sub-Antarctic islands | Manuele Bazzichetto, François Massol, Marta Carboni, Jonathan Lenoir, Jonas Johan Lembrechts, Rémi Joly, David Renault | <p>Aim Here, we aim to: (i) investigate the local effect of environmental and human-related factors on alien plant invasion in sub-Antarctic islands; (ii) explore the relationship between alien species features and their dependence on anthropogeni... |  | Biogeography, Biological invasions, Spatial ecology, Metacommunities & Metapopulations, Species distributions | Joaquín Hortal | | 2020-07-21 21:13:08 | View |
Intraspecific diversity loss in a predator species alters prey community structure and ecosystem functions
Allan Raffard, Julien Cucherousset, José M. Montoya, Murielle Richard, Samson Acoca-Pidolle, Camille Poésy, Alexandre Garreau, Frédéric Santoul, Simon Blanchet
https://doi.org/10.1101/2020.06.10.144337
Hidden diversity: how genetic richness affects species diversity and ecosystem processes in freshwater ponds
Recommended by Frederik De Laender based on reviews by Andrew Barnes and Jes Hines
Biodiversity loss can have important consequences for ecosystem functions, as exemplified by a large body of literature spanning at least three decades [1–3]. While connections between species diversity and ecosystem functions are now well-defined and understood, the importance of diversity within species is more elusive. Despite a surge in theoretical work on how intraspecific diversity can affect coexistence in simple community types [4,5], not much is known about how intraspecific diversity drives ecosystem processes in more complex community types. One particular challenge is that intraspecific diversity can be expressed as observable variation of functional traits, or instead subsist as genetic variation of which the consequences for ecosystem processes are harder to grasp.
Raffard et al. [6] examined how intraspecific biodiversity loss in a consumer fish changes species diversity at lower trophic levels and ecosystem processes in pond mesocosms. An interesting feature of this experiment is that it crosses functional and genetic intraspecific diversity. To do so, Raffard and colleagues measured and genotyped European minnow (P. phoxinus) individuals sampled from streams across southern France. Combining these collected specimens into experimental ponds allowed them to control functional (population variance of body size) and genetic intraspecific richness (number of genotypes).
Effects on minnow biomass production were mostly small; biomass was significantly reduced only when lowering both functional and genetic richness. However, the consequences for lower trophic levels (zooplankton and macroinvertebrates) were more pronounced and – importantly – not intuitive. For instance, the macroinvertebrate community was less species-diverse at higher minnow functional richness. If minnows with different body sizes would be the main regulator factors [7] explaining macroinvertebrate interactions, one would expect a more diverse set of minnow body sizes (i.e. higher functional minnow richness) to permit higher instead of lower macroinvertebrate richness. At the same time, the macroinvertebrate community was more species-diverse at higher minnow genotype richness, which could indicate unobserved minnow traits determining macroinvertebrate diversity more than the usual suspects (functional consumer richness). Such unobserved traits could be behavioral traits, allowing for resource partitioning among fish.
The consequences of functional minnow diversity loss on zooplankton diversity were negative, as expected in case body size differences among fish would facilitate coexistence of their zooplankton prey, as explained above. However, this was only the case when genetic diversity was high, suggesting nonstraightforward interactive effects of observed and non-observed traits on prey diversity.
The effects of functional and genetic minnow diversity loss on invertebrate (macroinvertebrates and zooplankton) abundance were more consistent than for invertebrate diversity. This suggests again nonstraightforward relationships in this experimental ecosystem, but now between invertebrate diversity and abundance. When using abundance as a proxy for an ecosystem process (which the authors did not), this result illustrates that biodiversity loss in multitrophic communities can have consequences that are challenging to interpret, let alone predict [8,9]. Path analyses showed how the observed changes of invertebrate diversity and abundance co-determined decomposition, a key ecosystem function. These path analyses had highest explanatory power show when including both kinds of intraspecific diversity.
Taken together, the results by Raffard and colleagues suggest that genetic consumer richness can drive species diversity of connected trophic levels and ecosystem processes with similar magnitude as functional diversity. Indeed, the effects of genetic consumer richness were shown to be so strong as to compensate or exacerbate the loss of observed functional richness. The exact mechanisms explaining these effects remain to be identified, however. The possibility that fish grazing by fish with different (observed or not observed) traits regulates coexistence among invertebrate prey, for instance, would depend on how strong fish consumption feeds back on prey growth during a 30-week experiment. As the authors indicate, detailed studies on resource partitioning among consumers (e.g. using stable isotope labelling) can shed light on these matters. Doing so may address a more fundamental question, which is if the mechanisms linking intraspecific diversity to function are different from those linking interspecific diversity to function, and at what time scales.
References
[1] Tilman D, Downing JA (1994) Biodiversity and stability in grasslands. Nature, 367, 363–365. https://doi.org/10.1038/367363a0
[2] Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, Venail P, Narwani A, Mace GM, Tilman D, Wardle DA, Kinzig AP, Daily GC, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S (2012) Biodiversity loss and its impact on humanity. Nature, 486, 59–67. https://doi.org/10.1038/nature11148
[3] De Laender F, Rohr JR, Ashauer R, Baird DJ, Berger U, Eisenhauer N, Grimm V, Hommen U, Maltby L, Meliàn CJ, Pomati F, Roessink I, Radchuk V, Brink PJV den (2016) Reintroducing Environmental Change Drivers in Biodiversity–Ecosystem Functioning Research. Trends in Ecology & Evolution, 31, 905–915. https://doi.org/10.1016/j.tree.2016.09.007
[4] Hart SP, Schreiber SJ, Levine JM (2016) How variation between individuals affects species coexistence. Ecology Letters, 19, 825–838. https://doi.org/10.1111/ele.12618
[5] Barabás G, D’Andrea R (2016) The effect of intraspecific variation and heritability on community pattern and robustness. Ecology Letters, 19, 977–986. https://doi.org/10.1111/ele.12636
[6] Raffard A, Cucherousset J, Montoya JM, Richard M, Acoca-Pidolle S, Poésy C, Garreau A, Santoul F, Blanchet S (2020) Intraspecific diversity loss in a predator species alters prey community structure and ecosystem functions. bioRxiv, 2020.06.10.144337, ver. 3 peer-reviewed and recommended by PCI Ecology. https://doi.org/10.1101/2020.06.10.144337
[7] Pásztor L, Botta-Dukát Z, Magyar G, Czárán T, Meszéna G. Theory-Based Ecology: A Darwinian approach. Oxford University Press. https://doi.org/10.1093/acprof:oso/9780199577859.001.0001
[8] Binzer A, Guill C, Rall BC, Brose U (2016) Interactive effects of warming, eutrophication and size structure: impacts on biodiversity and food-web structure. Global Change Biology, 22, 220–227. https://doi.org/10.1111/gcb.13086
[9] Schwarz B, Barnes AD, Thakur MP, Brose U, Ciobanu M, Reich PB, Rich RL, Rosenbaum B, Stefanski A, Eisenhauer N (2017) Warming alters energetic structure and function but not resilience of soil food webs. Nature Climate Change, 7, 895–900. https://doi.org/10.1038/s41558-017-0002-z
| Intraspecific diversity loss in a predator species alters prey community structure and ecosystem functions | Allan Raffard, Julien Cucherousset, José M. Montoya, Murielle Richard, Samson Acoca-Pidolle, Camille Poésy, Alexandre Garreau, Frédéric Santoul, Simon Blanchet | <p>Loss in intraspecific diversity can alter ecosystem functions, but the underlying mechanisms are still elusive, and intraspecific biodiversity-ecosystem function relationships (iBEF) have been restrained to primary producers. Here, we manipulat... |  | Community ecology, Ecosystem functioning, Experimental ecology, Food webs, Freshwater ecology | Frederik De Laender | Andrew Barnes | 2020-06-15 09:04:53 | View |
Uncertain predictions of species responses to perturbations lead to underestimate changes at ecosystem level in diverse systems
Recommended by Elisa Thebault based on reviews by Carlos Melian and 1 anonymous reviewer
Different sources of uncertainty are known to affect our ability to predict ecological dynamics (Petchey et al. 2015). However, the consequences of uncertainty on prediction biases have been less investigated, especially when predictions are scaled up to higher levels of organisation as is commonly done in ecology for instance. The study of Orr et al. (2020) addresses this issue. It shows that, in complex systems, the uncertainty of unbiased predictions at a lower level of organisation (e.g. species level) leads to a bias towards underestimation of change at higher level of organisation (e.g. ecosystem level). This bias is strengthened by larger uncertainty and by higher dimensionality of the system.
This general result has large implications for many fields of science, from economics to energy supply or demography. In ecology, as discussed in this study, these results imply that the uncertainty of predictions of species’ change increases the probability of underestimation of changes of diversity and stability at community and ecosystem levels, especially when species richness is high. The uncertainty of predictions of species’ change also increases the probability of underestimation of change when multiple ecosystem functions are considered at once, or when the combined effect of multiple stressors is considered.
The consequences of species diversity on ecosystem functions and stability have received considerable attention during the last decades (e.g. Cardinale et al. 2012, Kéfi et al. 2019). However, since the bias towards underestimation of change increases with species diversity, future studies will need to investigate how the general statistical effect outlined by Orr et al. might affect our understanding of the well-known relationships between species diversity and ecosystem functioning and stability in response to perturbations.
References
Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, Venail P, Narwani A, Mace GM, Tilman D, Wardle DA, Kinzig AP, Daily GC, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S (2012) Biodiversity loss and its impact on humanity. Nature, 486, 59–67. https://doi.org/10.1038/nature11148
Kéfi S, Domínguez‐García V, Donohue I, Fontaine C, Thébault E, Dakos V (2019) Advancing our understanding of ecological stability. Ecology Letters, 22, 1349–1356. https://doi.org/10.1111/ele.13340
Orr JA, Piggott JJ, Jackson A, Arnoldi J-F (2020) Why scaling up uncertain predictions to higher levels of organisation will underestimate change. bioRxiv, 2020.05.26.117200. https://doi.org/10.1101/2020.05.26.117200
Petchey OL, Pontarp M, Massie TM, Kéfi S, Ozgul A, Weilenmann M, Palamara GM, Altermatt F, Matthews B, Levine JM, Childs DZ, McGill BJ, Schaepman ME, Schmid B, Spaak P, Beckerman AP, Pennekamp F, Pearse IS (2015) The ecological forecast horizon, and examples of its uses and determinants. Ecology Letters, 18, 597–611. https://doi.org/10.1111/ele.12443
| Why scaling up uncertain predictions to higher levels of organisation will underestimate change | James Orr, Jeremy Piggott, Andrew Jackson, Jean-François Arnoldi | <p>Uncertainty is an irreducible part of predictive science, causing us to over- or underestimate the magnitude of change that a system of interest will face. In a reductionist approach, we may use predictions at the level of individual system com... |  | Community ecology, Ecosystem functioning, Theoretical ecology | Elisa Thebault | Anonymous | 2020-06-02 15:41:12 | View |
Insect herbivory on urban trees: Complementary effects of tree neighbours and predation
Alex Stemmelen, Alain Paquette, Marie-Lise Benot, Yasmine Kadiri, Hervé Jactel, Bastien Castagneyrol
https://doi.org/10.1101/2020.04.15.042317
Tree diversity is associated with reduced herbivory in urban forest
Recommended by Ruth Arabelle Hufbauer and Ian Pearse based on reviews by Ian Pearse and Freerk Molleman
Urban ecology, the study of ecological systems in our increasingly urbanized world, is crucial to planning and redesigning cities to enhance ecosystem services (Kremer et al. 2016), human health and well-being and further conservation goals (Dallimer et al. 2012). Urban trees are a crucial component of urban streets and parks that provide shade and cooling through evapotranspiration (Fung and Jim 2019), improve air quality (Lai and Kontokosta 2019), help control storm water (Johnson and Handel 2016), and conserve wildlife (Herrmann et al. 2012; de Andrade et al. 2020).
Ideally, management of urban forests strikes a balance between maintaining the health of urban trees while retaining those organisms, such as herbivores, that connect a tree to the urban ecosystem. Herbivory by arthropods can substantially affect tree growth and reproduction (Whittaker and Warrington 1985), and so understanding factors that influence herbivory in urban forests is important to effective management. At the same time, herbivorous arthropods are important as key components of urban bird diets (Airola and Greco 2019) and provide a backyard glimpse at forest ecosystems in an increasingly built environment (Pearse 2019). Maintenance of arthropod predators may be one way to retain arthropods in urban forests while keeping detrimental outbreaks of herbivores in check. In “Insect herbivory on urban trees: Complementary effects of tree neighbors and predation” Stemmelen and colleagues (Stemmelen et al. 2020) use a clever sampling design to show that insect herbivory decreases as the diversity of neighboring trees increased. By placing artificial larvae out on trees, they provide evidence that increased predation in higher diversity urban forest patches might drive patterns in herbivory. The paper also demonstrates the importance of tree species identity in determining leaf herbivory.
The implications of this research for urban foresters is that deliberately planting diverse urban forests will help manage insect herbivores and should thus improve tree health. Potential knock-on effects could be seen for the ecosystem services provided by urban forests. While it might be tempting to simply plant more of the species that are subject to low current rates of herbivory, other research on the long-term vulnerability of monocultures to attack by specialist pathogens and herbivores (Tooker and Frank 2012) cautions against such an approach. Furthermore, the importance of urban forest insects to birds, including migrating birds, argues for managing urban forests more holistically (Greco and Airola 2018).
Stemmelen et al. (2020) used an observational approach focused on urban forests in Montreal, Canada in their research. Their findings suggest follow-up research focused on a broader cross-section of urban forests across latitudes, as well as experimental research. Experiments could, for example, exclude avian predators with netting (e.g. (Marquis and Whelan 1994)) to evaluate the relative importance of birds to managing urban insects on trees, as well as the flip side of that equation, the important to birds of insects on urban trees.
In summary, Stemmelen and colleague’s manuscript illustrates clever sampling and use of observational data to infer broader ecological patterns. It is worth reading to better understand the role of diversity in driving plant-insect community interactions and given the implications of the findings for sustainable long-term management of urban forests.
References
Airola, D. and Greco, S. (2019). Birds and oaks in California’s urban forest. Int. Oaks, 30, 109–116.
de Andrade, A.C., Medeiros, S. and Chiarello, A.G. (2020). City sloths and marmosets in Atlantic forest fragments with contrasting levels of anthropogenic disturbance. Mammal Res., 65, 481–491. doi: https://doi.org/10.1007/s13364-020-00492-0
Dallimer, M., Irvine, K.N., Skinner, A.M.J., Davies, Z.G., Rouquette, J.R., Maltby, L.L., et al. (2012). Biodiversity and the Feel-Good Factor: Understanding Associations between Self-Reported Human Well-being and Species Richness. Bioscience, 62, 47–55. doi: https://doi.org/10.1525/bio.2012.62.1.9
Fung, C.K.W. and Jim, C.Y. (2019). Microclimatic resilience of subtropical woodlands and urban-forest benefits. Urban For. Urban Green., 42, 100–112. doi: https://doi.org/10.1016/j.ufug.2019.05.014
Greco, S.E. and Airola, D.A. (2018). The importance of native valley oaks (Quercus lobata) as stopover habitat for migratory songbirds in urban Sacramento, California, USA. Urban For. Urban Green., 29, 303–311. doi: https://doi.org/10.1016/j.ufug.2018.01.005
Herrmann, D.L., Pearse, I.S. and Baty, J.H. (2012). Drivers of specialist herbivore diversity across 10 cities. Landsc. Urban Plan., 108, 123–130. doi: https://doi.org/10.1016/j.landurbplan.2012.08.007
Johnson, L.R. and Handel, S.N. (2016). Restoration treatments in urban park forests drive long-term changes in vegetation trajectories. Ecol. Appl., 26, 940–956. doi: https://doi.org/10.1890/14-2063
Kremer, P., Hamstead, Z., Haase, D., McPhearson, T., Frantzeskaki, N., Andersson, E., et al. (2016). Key insights for the future of urban ecosystem services research. Ecol. Soc., 21: 29. doi: http://doi.org/10.5751/ES-08445-210229
Lai, Y. and Kontokosta, C.E. (2019). The impact of urban street tree species on air quality and respiratory illness: A spatial analysis of large-scale, high-resolution urban data. Heal. Place, 56, 80–87. doi: https://doi.org/10.1016/j.healthplace.2019.01.016
Marquis, R.J. and Whelan, C.J. (1994). Insectivorous birds increase growth of white oak through consumption of leaf-chewing insects. Ecology, 75, 2007–2014. doi: https://doi.org/10.2307/1941605
Pearse, I.S. (2019). Insect herbivores on urban native oak trees. Int. Oaks, 30, 101–108.
Stemmelen, A., Paquette, A., Benot, M.-L., Kadiri, Y., Jactel, H. and Castagneyrol, B. (2020) Insect herbivory on urban trees: Complementary effects of tree neighbours and predation. bioRxiv, 2020.04.15.042317, ver. 5 peer-reviewed and recommended by PCI Ecology. doi: https://doi.org/10.1101/2020.04.15.042317
Tooker, J. F., and Frank, S. D. (2012). Genotypically diverse cultivar mixtures for insect pest management and increased crop yields. J. Appl. Ecol., 49(5), 974-985. doi: https://doi.org/10.1111/j.1365-2664.2012.02173.x
Whittaker, J.B. and Warrington, S. (1985). An experimental field study of different levels of insect herbivory induced By Formica rufa predation on Sycamore (Acer pseudoplatanus) III. Effects on Tree Growth. J. Appl. Ecol., 22, 797. doi: https://doi.org/10.2307/2403230
| Insect herbivory on urban trees: Complementary effects of tree neighbours and predation | Alex Stemmelen, Alain Paquette, Marie-Lise Benot, Yasmine Kadiri, Hervé Jactel, Bastien Castagneyrol | <p>Insect herbivory is an important component of forest ecosystems functioning and can affect tree growth and survival. Tree diversity is known to influence insect herbivory in natural forest, with most studies reporting a decrease in herbivory wi... |  | Biodiversity, Biological control, Community ecology, Ecosystem functioning, Herbivory | Ruth Arabelle Hufbauer | | 2020-04-20 13:49:36 | View |
The role of behavior and habitat availability on species geographic expansion
Recommended by Esther Sebastián González based on reviews by Caroline Marie Jeanne Yvonne Nieberding, Pizza Ka Yee Chow, Tim Parker and 1 anonymous reviewer
Understanding the relative importance of species-specific traits and environmental factors in modulating species distributions is an intriguing question in ecology [1]. Both behavioral flexibility (i.e., the ability to change the behavior in changing circumstances) and habitat availability are known to influence the ability of a species to expand its geographic range [2,3]. However, the role of each factor is context and species dependent and more information is needed to understand how these two factors interact. In this pre-registration, Logan et al. [4] explain how they will use Great-tailed grackles (Quiscalus mexicanus), a species with a flexible behavior and a rapid geographic range expansion, to evaluate the relative role of habitat and behavior as drivers of the species’ expansion [4]. The authors present very clear hypotheses, predicted results and also include alternative predictions. The rationales for all the hypotheses are clearly stated, and the methodology (data and analyses plans) are described with detail. The large amount of information already collected by the authors for the studied species during previous projects warrants the success of this study. It is also remarkable that the authors will make all their data available in a public repository, and that the pre-registration in already stored in GitHub, supporting open access and reproducible science. I agree with the three reviewers of this pre-registration about its value and I think its quality has largely improved during the review process. Thus, I am happy to recommend it and I am looking forward to seeing the results.
References
[1] Gaston KJ. 2003. The structure and dynamics of geographic ranges. Oxford series in Ecology and Evolution. Oxford University Press, New York.
[2] Sol D, Lefebvre L. 2000. Behavioural flexibility predicts invasion success in birds introduced to new zealand. Oikos. 90(3): 599–605. https://doi.org/10.1034/j.1600-0706.2000.900317.x
[3] Hanski I, Gilpin M. 1991. Metapopulation dynamics: Brief history and conceptual domain. Biological journal of the Linnean Society. 42(1-2): 3–16. https://doi.org/10.1111/j.1095-8312.1991.tb00548.x
[4] Logan CJ, McCune KB, Chen N, Lukas D. 2020. Implementing a rapid geographic range expansion - the role of behavior and habitat changes (http://corinalogan.com/Preregistrations/gxpopbehaviorhabitat.html) In principle acceptance by PCI Ecology of the version on 16 Dec 2021 https://github.com/corinalogan/grackles/blob/0fb956040a34986902a384a1d8355de65010effd/Files/Preregistrations/gxpopbehaviorhabitat.Rmd.
| Implementing a rapid geographic range expansion - the role of behavior and habitat changes | Logan CJ, McCune KB, Chen N, Lukas D | <p>It is generally thought that behavioral flexibility, the ability to change behavior when circumstances change, plays an important role in the ability of a species to rapidly expand their geographic range (e.g., Lefebvre et al. (1997), Griffin a... |  | Behaviour & Ethology, Biological invasions, Dispersal & Migration, Foraging, Habitat selection, Human impact, Phenotypic plasticity, Preregistrations, Zoology | Esther Sebastián González | Anonymous, Caroline Marie Jeanne Yvonne Nieberding, Tim Parker | 2020-05-14 11:18:57 | View |
Explore and move: a key to success in a changing world?
Recommended by Blandine Doligez based on reviews by Joe Nocera, Marion Nicolaus and Laure Cauchard
Changes in the spatial range of many species are one of the major consequences of the profound alteration of environmental conditions due to human activities. Some species expand, sometimes spectacularly during invasions; others decline; some shift. Because these changes result in local biodiversity loss (whether local species go extinct or are replaced by colonizing ones), understanding the factors driving spatial range dynamics appears crucial to predict biodiversity dynamics. Identifying the factors that shape individual movement is a main step towards such understanding. The study described in this preregistration (McCune et al. 2020) falls within this context by testing possible links between individual exploration behaviour and movements related to daily space use in an avian study model currently rapidly expanding, the great-tailed grackle (Quiscalus mexicanus).
Movement and exploration: which direction(s) for the link between exploration and dispersal?
Individuals are known to differ in their tendency to explore the environment (Réale et al. 2007; Wolf and Weissing 2012) and therefore in their motivation to move. Accordingly, exploration has been shown to relate to dispersal behaviour, i.e. movements between breeding sites (Dingemanse et al. 2003, Le Galliard et al. 2011, Rasmussen and Belk 2012; reviews in Cote et al. 2010, Ronce et al. 2012). Yet, the mechanisms underlying this link often remain unclear, due to the correlative nature of the data. A classical assumption is that dispersers may benefit from a high capacity to explore, allowing them to familiarize quicker with their new environment once reached, thus alleviating dispersal costs (Bonte et al. 2012). The association between dispersal and exploration would in this case result from selection for this combination of traits (Ronce et al. 2012), even though dispersal event itself may be independent from (and precede the effect of) exploration behaviour. Alternatively (but not exclusively), dispersal may simply be the final outcome of longer movements by individuals exploring larger ranges (Badyaev et al. 1996, Schliehe-Diecks et al. 2012). In the absence of easy ways to manipulate dispersal behaviour, on the one hand, and exploration tendency, on the other hand, investigating detailed, small-scale individual movements in relation to exploration should thus shed light on which processes may yield the observed relations between exploration as an individual personality trait and large-scale, long-term movements, such as dispersal, underlying species range dynamics.
In this project, the exploration behaviour of grackles will be measured in controlled conditions using standardized tests in captivity (McCune et al. 2019) before individuals are released and their daily space use behaviour will then be measured using remote tracking over long time periods (McCune et al. 2020). Importantly, these coupled measures will be obtained for individuals captured in three different populations: within the historical range of the species, in the middle of its expanding range and at the edge of the range (McCune et al. 2020). Therefore, the project will test (i) whether daily space use of individuals is linked to their intrinsic exploration tendency and (ii) whether space use differs between individuals from different populations along the expanding range. The preregistration echoes a complementary project by the same team that will focus on exploration and test (iii) whether exploration tendency differs between individuals from these different populations. Taken together, these three analyses will therefore provide solid background information to assess the role of exploration in the individuals’ decisions leading to movement and range dynamics in this species.
As underlined in the preregistration, previous studies addressing the links between individual exploration behaviour and movements have mostly focused on dispersal. A first type of studies have (as will be done here) measured exploration behaviour of individuals, often in captivity (Dingemanse et al. 2003, Korsten et al. 2013) but also in the wild (Rasmussen and Belk 2012, Debeffe et al. 2013), and related these measures to subsequent dispersal behaviour. The (often implicit) underlying assumption is that more exploratory individuals will be more likely to move further, explore different habitats and thus end up breeding farther than less explorative ones. In other words, exploration tendency precedes and drives dispersal. Sometimes, exploratory behaviour is measured on individuals of known dispersal status, i.e. after the dispersal event (Hoset et al. 2011), in which case selection for certain exploration phenotypes among dispersers may already have occurred. Besides this first approach, another type of studies have measured ‘exploration’ behaviour under the form of prospecting movements of individuals and linked these movements to subsequent dispersal (often in the context of habitat selection). While these studies were in the past based on direct thus potentially biased observations (Reed et al. 1999), they now rely more and more on technological advances using (miniaturized) remote tracking devices (Ponchon et al. 2013) that provide far more complete and unbiased movement data, and sometimes also complementary measures of individuals’ internal state. In this case, the implicit assumption is that individuals prospecting farther and/or in more habitat patches will be more likely to settle in a site located farther away from their departure site, because of a more exhaustive sampling of possible sites allowing individuals to identify higher-quality sites (Badyaev et al. 1996). In other words, exploration tendency would not directly lead to higher movements or longer distances, but would allow individuals to optimize their habitat choice among more numerous options, thus leading to an increased dispersal probability or distance; the relation between exploration and dispersal would thus be indirect. Prospecting studies address more closely the underlying mechanisms of movement; however, they cannot easily separate intrinsic individual exploratory tendency from the prospecting movements themselves, with potential feedback effects of the information already gathered on future exploration of other sites or patches, thus on subsequent movements.
By focusing on individual daily space use movements as a mechanistic approach to understand large-scale movements potentially involved in colonization and range expansion, the grackle study described in this preregistration (McCune et al. 2020) will thus contribute to bridge the knowledge gaps between exploration and dispersal. By linking exploration measures obtained from a battery of standardized tests conducted in controlled conditions to individual daily space use and movements recorded in the wild, the grackle project is set in between previous studies addressing the links between exploration and dispersal: it will document exploration in a separate and independent context with respect to the movements themselves, and it will use a mechanistic view of detailed movements by the same individuals in the wild to explore potential implications for dispersal and range expansion. Testing differences between the three study populations over the species range will indeed inform about potential large-scale, population implications of among-individual variation in the link between exploration and movements. Because this study will only measure already settled adult individuals whose previous history is unknown, there will nevertheless be no direct possible exploration of the link with either previous or subsequent dispersal behaviour. Thus, the potential links studied here relate more directly to post-dispersal benefits of exploration for an optimal exploitation of the new environment. Yet, if exploration is a life-long personality trait linked to daily movement patterns, it may also relate to natal dispersal movements in young individuals.
Evolutionary and conservation perspectives
If the results of the project reveal that exploration tendency and daily space use movements are indeed linked, and that individuals from populations across the species range differ in these traits, new questions will emerge. A first question would be whether such among-individual differences are at the origin of range expansion or rather one of its consequences since, again, we deal with correlative data here. In other words, individuals may differ in exploration tendency, and this may confer them different ability to move around, find and colonize new habitats; or individuals may show differences in exploration following arrival in a new habitat, either because more explorative individuals gain fitness benefits and are thus selected, or because of behavioural plasticity and post-colonization adjustment of exploration behaviour when facing new ecological and social conditions in the new environment. Another open question relates to the link between daily space use and dispersal: is dispersal a by-product of higher daily movements that allow individuals to discover new favorable places where to settle? Exploring this link could involve measuring just fledged individuals before natal dispersal occurs and/or individuals chosen according to their own dispersal history, and this would then imply long-term population monitoring as an efficient (but constraining) tool to address such questions. Finally, assessing the fitness consequences of the link between exploration and space use behaviour, and whether these consequences differ between populations along the range expansion, would also be needed to understand the contribution of this link to the invasion success of this species.
The study model chosen for this project is a rapidly expanding species. Importantly, however, and as emphasized in the preregistration, documenting links between exploration and daily space use patterns as well as differences between populations with different trajectories can provide crucial information in general to understand population persistence in response to global climate and landscape changes, both regarding invasion ability or extinction risk. The information should be key to assess the probability that a species may decline, persist or expand in studies addressing biodiversity and community dynamics in a changing world.
References
Badayev, A. V., Martin, T. E and Etges, W. J. 1996. Habitat sampling and habitat selection by female wild turkeys: ecological correlates and reproductive consequences. Auk 113: 636-646. doi: https://doi.org/10.2307/4088984
Bonte, D. et al. 2012. Costs of dispersal. Biological Reviews 87: 290-312. doi: https://doi.org/10.1111/j.1469-185X.2011.00201.x
Cote, J., Clobert, J., Brodin, T., Fogarty, S. and Sih, A. 2010. Personality-dependent dispersal: characterization, ontogeny and consequences for spatially structured populations. Philosophical Transactions of the Royal Society B 365: 4065-4576. doi: https://doi.org/10.1098/rstb.2010.0176
Debeffe, L., Morellet, N., Cargnelutti, B., Lourtet, B., Coulon, A., Gaillard, J.-M., Bon, R. and Hewison A. J. M. 2013. Exploration as a key component of natal dispersal: dispersers explore more than philopatric individuals in roe deer. Animal Behaviour 86: 143-151. doi: https://doi.org/10.1016/j.anbehav.2013.05.005
Dingemanse, N. J., Both, C., van Noordwijk, A. J., Rutten, A. L. and Drent, P. J. 2003. Natal dispersal and personalities in great tits (Parus major). Proceedings of the Royal Society B 270: 741-747. doi: https://doi.org/10.1098/rspb.2002.2300
Hoset, K. S., Ferchaud, A.-L., Dufour, F., Mersch, D., Cote, J. and Le Galliard, J.-F. 2011. Natal dispersal correlates with behavioral traits that are not consistent across early life stages. Behavioral Ecology 22: 176–183. doi: https://doi.org/10.1093/beheco/arq188
Korsten, P., van Overveld, T., Adriaensen, F. and Matthysen, E. 2013. Genetic integration of local dispersal and exploratory behaviour in a wild bird. Nature Communications 4: 2362. doi: https://doi.org/10.1038/ncomms3362
Le Galliard, J.-F., Rémy, A., Ims, R. A. and Lambin, X. 2011. Patterns and processes of dispersal behaviour in arvicoline rodents. Molecular Ecology 21: 505-523. doi: https://doi.org/10.1111/j.1365-294X.2011.05410.x
McCune K, Ross C, Folsom M, Bergeron L, Logan CJ. 2020. Does space use behavior relate to exploration in a species that is rapidly expanding its geographic range? http://corinalogan.com/Preregistrations/gspaceuse.html In principle acceptance by PCI Ecology of the version on 23 Sep 2020 https://github.com/corinalogan/grackles/blob/master/Files/Preregistrations/gspaceuse.Rmd.
McCune K, MacPherson M, Rowney C, Bergeron L, Folsom M, Logan CJ. 2019. Is behavioral flexibility linked with exploration, but not boldness, persistence, or motor diversity? (http://corinalogan.com/Preregistrations/gexploration.html) In principle acceptance by PCI Ecology of the version on 27 Mar 2019 https://github.com/corinalogan/grackles/blob/master/Files/Preregistrations/gexploration.Rmd
Ponchon, A., Grémillet, D., Doligez, B., Chambert, T., Tveraa, T., González-Solís, J. and Boulinier, T. 2013. Tracking prospecting movements involved in breeding habitat selection: insights, pitfalls and perspectives. Methods in Ecology and Evolution 4: 143-150. doi: https://doi.org/10.1111/j.2041-210x.2012.00259.x
Rasmussen, J. E. and Belk, M. C. 2012. Dispersal behavior correlates with personality of a North American fish. Current Zoology 58: 260–270. doi: https://doi.org/10.1093/CZOOLO%2F58.2.260
Réale, D., Reader, S. M., Sol, D., McDougall, P. T. and Dingemanse, N. J. 2007. Integrating animal temperament within ecology and evolution. Biological Reviews 82: 291-318. doi: https://doi.org/10.1111/j.1469-185x.2007.00010.x
Reed, J. M., Boulinier, T., Danchin, E. and Oring, L. W. 1999. Informed dispersal: prospecting by birds for breeding sites. Current Ornithology 15: 189-259. doi: https://doi.org/10.1007/978-1-4757-4901-4_5
Ronce, O. and Clobert, J. 2012. Dispersal syndromes. pp. 119-138 In Dispersal Ecology and Evolution (eds. Clobert, J., Baguette, M., Benton, T. G. and Bullock, J. M.), pp. 119-138. Oxford University Press.
Schliehe-Diecks, S., Eberle, M. and Kappeler, P. M. 2012. Walk the line - dispersal movements of gray mouse lemurs (Microcebus murinus). Behavioral Ecology and Sociobiology 66: 1175-1185. doi: https://dx.doi.org/10.1007%2Fs00265-012-1371-y
Wolf, M. and Weissing, F. J. 2012. Animal personalities: consequences for ecology and evolution. Trends in Ecology and Evolution 27: 452-461. doi: https://doi.org/10.1016/j.tree.2012.05.001
| Does space use behavior relate to exploration in a species that is rapidly expanding its geographic range? | Kelsey B. McCune, Cody Ross, Melissa Folsom, Luisa Bergeron, Corina Logan | Great-tailed grackles (Quiscalus mexicanus) are rapidly expanding their geographic range (Wehtje 2003). Range expansion could be facilitated by consistent behavioural differences between individuals on the range edge and those in other parts of th... |  | Behaviour & Ethology, Biological invasions, Conservation biology, Habitat selection, Phenotypic plasticity, Preregistrations, Spatial ecology, Metacommunities & Metapopulations | Blandine Doligez | | 2019-09-30 19:27:40 | View |
How citizen science could improve Species Distribution Models and their independent assessment
Florence Matutini, Jacques Baudry, Guillaume Pain, Morgane Sineau, Josephine Pithon
https://doi.org/10.1101/2020.06.02.129536
Citizen science contributes to SDM validation
Recommended by Francisco Lloret based on reviews by Maria Angeles Perez-Navarro and 1 anonymous reviewer
Citizen science is becoming an important piece for the acquisition of scientific knowledge in the fields of natural sciences, and particularly in the inventory and monitoring of biodiversity (McKinley et al. 2017). The information generated with the collaboration of citizens has an evident importance in conservation, by providing information on the state of populations and habitats, helping in mitigation and restoration actions, and very importantly contributing to involve society in conservation (Brown and Williams 2019).
An obvious advantage of these initiatives is the ability to mobilize human resources on a large territorial scale and in the medium term, which would otherwise be difficult to finance. The resulting increasing information then can be processed with advanced computational techniques (Hochachka et al 2012; Kelling et al. 2015), thus improving our interpretation of the distribution of species. Specifically, the ability to obtain information on a large territorial scale can be integrated into studies based on Species Distribution Models SDMs. One of the common problems with SDMs is that they often work from species occurrences that have been opportunistically recorded, either by professionals or amateurs. A great challenge for data obtained from non-professional citizens, however, remains to ensure its standardization and quality (Kosmala et al. 2016). This requires a clear and effective design, solid volunteer training, and a high level of coordination that turns out to be complex (Brown and Williams 2019). Finally, it is essential to perform a quality validation following scientifically recognized standards, since they are often conditioned by errors and biases in obtaining information (Bird et al. 2014). There are two basic approaches to obtain the necessary data for this validation: getting it from an external source (external validation), or allocating a part of the database itself (internal validation or cross-validation) to this function.
Matutini et al. (2020) in his work 'How citizen science could improve Species Distribution Models and their independent assessment' shows a novel application of the data generated by a citizen science initiative ('Un Dragon dans mon Jardin') by providing an external source for the validation of SDMs, as a tool to construct habitat suitability maps for nine species of amphibians in western France. Importantly, 'Un Dragon dans mon Jardin' contains standardized presence-absence data, the approximation recognized as the most robust (Guisan, et al. 2017). The SDMs to be validated, in turn, were based on opportunistic information obtained by citizens and professionals. The result shows the usefulness of this external data source by minimizing the overestimation of model accuracy that is obtained with cross-validation with the internal evaluation dataset. It also shows the importance of properly filtering the information obtained by citizens by determining the threshold of sampling effort.
The destiny of citizen science is to be integrated into the complex world of science. Supported by the increasing level of the formation of society, it is becoming a fundamental piece in the scientific system dedicated to the study of biodiversity and its conservation. After funding for scientists specialized in the recognition of biodiversity has been cut back, we are seeing a transformation of the activity of these scientists towards the design, coordination, training and verification of programs for the acquisition of field information obtained by citizens. A main goal is that a substantial part of this information will eventually get integrated into the scientific system, and rigorous verification process a fundamental element for such purpose, as shown by Matutini et al. (2020) work.
References
[1] Bird TJ et al. (2014) Statistical solutions for error and bias in global citizen science datasets. Biological Conservation 173: 144-154. doi: 10.1016/j.biocon.2013.07.037
[2] Brown ED and Williams BK (2019) The potential for citizen science to produce reliable and useful information in ecology. Conservation Biology 33: 561-569. doi: 10.1111/cobi.13223
[3] Guisan A, Thuiller W and Zimmermann N E (2017) Habitat Suitability and Distribution Models: With Applications in R. The University of Chicago Press. doi: 10.1017/9781139028271
[4] Hochachka WM, Fink D, Hutchinson RA, Sheldon D, Wong WK and Kelling S (2012) Data-intensive science applied to broad-scale citizen science. Trens Ecol Evol 27: 130-137. doi: 10.1016/j.tree.2011.11.006
[5] Kelling S, Fink D, La Sorte FA, Johnston A, Bruns NE and Hochachka WM (2015) Taking a ‘Big Data’ approach to data quality in a citizen science project. Ambio 44(Supple. 4):S601-S611. doi: 10.1007/s13280-015-0710-4
[6] Kosmala M, Wiggins A, Swanson A and Simmons B (2016) Assessing data quality in citizen science. Front Ecol Environ 14: 551–560. doi: 10.1002/fee.1436
[7] Matutini F, Baudry J, Pain G, Sineau M and Pithon J (2020) How citizen science could improve Species Distribution Models and their independent assessment. bioRxiv, 2020.06.02.129536, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/2020.06.02.129536
[8] McKinley DC et al. (2017) Citizen science can improve conservation science, natural resource management, and environmental protection. Biological Conservation 208:15-28. doi: 10.1016/j.biocon.2016.05.015
| How citizen science could improve Species Distribution Models and their independent assessment | Florence Matutini, Jacques Baudry, Guillaume Pain, Morgane Sineau, Josephine Pithon | <p>Species distribution models (SDM) have been increasingly developed in recent years but their validity is questioned. Their assessment can be improved by the use of independent data but this can be difficult to obtain and prohibitive to collect.... |  | Biodiversity, Biogeography, Conservation biology, Habitat selection, Spatial ecology, Metacommunities & Metapopulations, Species distributions, Statistical ecology | Francisco Lloret | | 2020-06-03 09:36:34 | View |
The dynamics of spawning acts by a semelparous fish and its associated energetic costs
Cédric Tentelier, Colin Bouchard, Anaïs Bernardin, Amandine Tauzin, Jean-Christophe Aymes, Frédéric Lange, Charlotte Recapet, Jacques Rives
https://doi.org/10.1101/436295
Extreme weight loss: when accelerometer could reveal reproductive investment in a semelparous fish species
Recommended by Francois-Xavier Dechaume-Moncharmont based on reviews by Aidan Jonathan Mark Hewison, Loïc Teulier and 1 anonymous reviewer
Continuous observation of animal behaviour could be quite a challenge in the field, and the situation becomes even more complicated with aquatic species mostly active at night. In such cases, biologging techniques are real game changers in ecology, behavioural ecology or eco-physiology. An accelerating number of methodological applications of these tools in natural condition are thus published each year [1]. Biologging is not limited to movement ecology. For instance, fine grain information about energy expenditure can be inferred from body acceleration [2], and accelerometers has already proven useful in monitoring reproductive costs in some fish species [3,4]. The first part of the study by Tentelier et al. [5] is in line with this growing literature. It describes measurements of energy expenditure during reproduction in a fish species, Allis shad (Alosa Alosa), based on tail beat frequency and occurrence of spawning acts. The study has been convincingly conducted, and the results are important for fish biologists. But this is not the whole story: the authors added to this otherwise classical study a very original and insightful analysis which deserves closer interest.
Tentelier et al. propose to use static accelerometer to monitor change in body roundness through the reproductive season. These semelparous fish first mature and built up reserves in the Atlantic Ocean and migrate into fresh water to reproduce. Contrary to iteroparous species, female shads do not have to strategically preserve energy for future reproduction. The females die few days after spawning having exhausted their energetic reserves: they typically lose almost half of their body mass during the spawning season. The beautiful idea in this study was to track down information about this dramatic slimming in the accelerometer data. Indeed, the accelerometer was attached on the side of the fish (close to the dorsal fin). A change in its angle with the vertical plane could be correlated with the change in roundness, the angle declining with the female thinning. Accelerometers have already been used to record body posture [6] but, in the present study, the novelty was to monitor the change in body shape.
Unfortunately, the data by Tentelier et al. are inconclusive so far. Broadly speaking, the accelerometer angle recorded declined through the spawning season, indicating an average slimming of the females, but there was no correlation between the change in angle and the mass loss at the individual level. This was partly due to the fact that the dorsal position of the accelerometer was not optimized to measures egg laying whose effects are mostly observable on ventral side.
Yet, this nice idea deserves more scrutiny. The method seems to be sensitive enough to detect inflation of swim bladder, the gas-filled organ helping the fish to control their position in the water column, as the accelerometer angle increased when the fish stayed close to the water surface. Additional works and proper calibration are certainly needed to validate the use of accelerometer angle as a proxy for body roundness. The actual data were not strong enough to justify a standalone publication on the subject, but it would have been shame to lose traces of such analysis and keep it in the file drawer. This is why I strongly support its report as a side question in a broader study. Science progresses not only with neat conclusive studies but also when unexpected (apparently anecdotal) observations stimulate new researches.
References
[1] Börger L, Bijleveld AI, Fayet AL, Machovsky‐Capuska GE, Patrick SC, Street GM and Vander Wal E. (2020) Biologging special feature. J. Anim. Ecol. 89, 6–15. 10.1111/1365-2656.13163
[2] Wilson RP et al. (2020) Estimates for energy expenditure in free‐living animals using acceleration proxies: A reappraisal. J. Anim. Ecol. 89, 161–172. 10.1111/1365-2656.13040
[3] Tsuda Y, Kawabe R, Tanaka H, Mitsunaga Y, Hiraishi T, Yamamoto K and Nashimoto K. (2006) Monitoring the spawning behaviour of chum salmon with an acceleration data logger. Ecol. Freshw. Fish 15, 264–274. 10.1111/j.1600-0633.2006.00147.x
[4] Sakaji H, Hamada K, Naito Y. 2018 Identifying spawning events of greater amberjack using accelerometers. Mar. Biol. Res. 14, 637–641. 10.1080/17451000.2018.1492140
[5] Tentelier C, Bouchard C, Bernardin A, Tauzin A, Aymes J-C, Lange F, Récapet C, Rives J (2020) The dynamics of spawning acts by a semelparous fish and its associated energetic costs. bioRxiv, 436295. doi: 10.1101/436295 ver. 7 peer-reviewed and recommended by PCI Ecology. 10.1101/436295
[6] Brown DD, Kays R, Wikelski M, Wilson R, Klimley AP. 2013 Observing the unwatchable through acceleration logging of animal behavior. Anim. Biotelemetry 1, 20. 10.1186/2050-3385-1-20
| The dynamics of spawning acts by a semelparous fish and its associated energetic costs | Cédric Tentelier, Colin Bouchard, Anaïs Bernardin, Amandine Tauzin, Jean-Christophe Aymes, Frédéric Lange, Charlotte Recapet, Jacques Rives | <p>1. During the reproductive season, animals have to manage both their energetic budget and gamete stock. In particular, for semelparous capital breeders with determinate fecundity and no parental care other than gametic investment, the depletion... |  | Behaviour & Ethology, Freshwater ecology, Life history | Francois-Xavier Dechaume-Moncharmont | | 2020-06-04 15:18:56 | View |





