Data-based, synthesis-driven: setting the agenda for computational ecology
Timothée Poisot, Richard Labrie, Erin Larson, Anastasia Rahlin
10.1101/150128
Some thoughts on computational ecology from people who I’m sure use different passwords for each of their accounts
Recommended by Phillip P.A. Staniczenko based on reviews by Matthieu Barbier and 1 anonymous reviewer
Are you an ecologist who uses a computer or know someone that does?
Even if your research doesn’t rely heavily on advanced computational techniques, it likely hasn’t escaped your attention that computers are increasingly being used to analyse field data and make predictions about the consequences of environmental change. So before artificial intelligence and robots take over from scientists, now is great time to read about how experts think computers could make your life easier and lead to innovations in ecological research.
In “Data-based, synthesis-driven: setting the agenda for computational ecology”, Poisot and colleagues [1] provide a brief history of computational ecology and offer their thoughts on how computational thinking can help to bridge different types of ecological knowledge.
In this wide-ranging article, the authors share practical strategies for realising three main goals: (i) tighter integration of data and models to make predictions that motivate action by practitioners and policy-makers; (ii) closer interaction between data-collectors and data-users; and (iii) enthusiasm and aptitude for computational techniques in future generations of ecologists.
The key, Poisot and colleagues argue, is for ecologists to “engage in meaningful dialogue across disciplines, and recognize the currencies of their collaborations.”
Yes, this is easier said than done. However, the journey is much easier with a guide and when everyone involved serves to benefit not only from the eventual outcome, but also the process.
References
[1] Poisot, T., Labrie, R., Larson, E., & Rahlin, A. (2018). Data-based, synthesis-driven: setting the agenda for computational ecology. BioRxiv, 150128, ver. 4 recommended and peer-reviewed by PCI Ecology. doi: 10.1101/150128
| Data-based, synthesis-driven: setting the agenda for computational ecology | Timothée Poisot, Richard Labrie, Erin Larson, Anastasia Rahlin | Computational ecology, defined as the application of computational thinking to ecological problems, has the potential to transform the way ecologists think about the integration of data and models. As the practice is gaining prominence as a way to... | 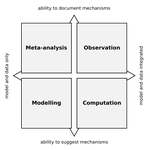 | Meta-analyses, Statistical ecology, Theoretical ecology | Phillip P.A. Staniczenko | | 2018-02-05 20:51:41 | View |
Touchy matter: the delicate balance between Morgan’s canon and open-minded description of advanced cognitive skills in the animal
Recommended by Francois-Xavier Dechaume-Moncharmont based on reviews by Valérie Dufour and Alex Taylor based on reviews by Valérie Dufour and Alex Taylor
In a recent paper published in PNAS, Fayet et al. [1] reported scarce field observations of two Atlantic puffins (four years apart) apparently scratching their bodies using sticks, which was interpreted by the authors as evidence of tool use in this species. In a short response, Benjamin Farrar [2] raises serious concerns about this interpretation and proposes simpler, more parsimonious, mechanisms explaining the observed behaviour: a textbook case of Morgan's canon.
In virtually all introductory lectures on animal behaviour, students are advised to exercise caution when interpreting empirical data and weighting alternative explanations. We are sometimes prisoner of our assumptions: our desire of beliefs in advanced cognitive skills in non-human species make us more receptive to facts confirming our preconceptions than to simpler, less exciting, interpretations (a phenomenon known as "confirmation bias" in psychology). We must resist the temptation to accept appealing explanations without enough critical thinking. Our students are thus taught to apply the Lloyd Morgan's canon, a variant of one of the most important heuristics in Science, the principle of parsimony or Occam's razor, rephrased by Morgan [3, page 53] in the context of animal behaviour: "In no case may we interpret an action as the outcome of a higher psychical faculty, if it can be interpreted as the outcome of the exercise of one that stands lower in the psychological scale". In absence of evidence to the contrary, one should postulate the simplest cognitive skill consistent with the observed behaviour. While sometimes criticized from an epistemological point of view [4-6], it remains an essential and largely accepted framework of animal cognition. It has repeatedly proved to be a useful guide in the minefield of comparative psychology. Classical ethology questions related to the existence of, for instance, meta-cognition [7], intentionality or problem solving [8] have been convincingly investigated using this principle.
Yet, there is a downside to this conservative approach. Blind reference to Morgan's canon may narrow our theoretical thinking about animal cognition [7,9]. It could be counter-productive to systematically deny advanced cognitive skills in animals. On the contrary, keeping our mind open to unplanned observations, unexpected discoveries, or serendipity [10], and being prepared to accept new hypotheses, sometimes fairly remote from the dominant paradigm, may be a fruitful research strategy. To quote Darwin's famous letter to Alfred Wallace: "I am a firm believer, that without speculation there is no good and original observation" [11]. Brief notes in specialized scientific journals, or even in grey literature (by enthusiast amateur ornithologists, ichthyologists, or entomologists), constitutes a rich array of anecdotal observations. For instance, Sol et al. [12] convincingly compared the innovation propensity across bird species by screening ornithology literature using keywords like 'never reported', 'not seen before', 'first report', 'unusual' or 'novel'. Even if "the plural of anecdote is not data" as the saying goes, such descriptions of novel behaviours, even single-subject observations, are indisputably precious: taxonomic ubiquity of a behaviour is a powerful argument in favour of evolutionary convergence. Of course, a race to the bottom, amplified by the inevitable media hypes around scientific articles questioning human exceptionalism, is another possible scientific trap for behavioural biologists in search of skills characteristic of so-called advanced species, but never described so far in supposedly cognitively simpler organisms. As stated by Franz de Waal [9]: "I have nothing against anecdotes, especially if they have been caught on camera or come from reputable observers who know their animals; but I do view them as a starting point of research, never an end point".
In the case of the two video observations of puffins apparently using sticks as scratching tool, it must be considered as a mere anecdote unless scientists systematically investigate this behaviour. In his constructive criticism of Fayet et al.'s paper, Benjamin Farrar [2] proposes interesting directions of research and testable predictions. A correlation between the background rate of stick picking and the rate of stick preening would indicate that this behaviour was more likely explained by fluke than genuine innovation in this species.
References
[1] Fayet, A. L., Hansen, E. S., and Biro, D. (2020). Evidence of tool use in a seabird. Proceedings of the National Academy of Sciences, 117(3), 1277–1279. doi: 10.1073/pnas.1918060117
[2] Farrar, B. G. (2020). Evidence of tool use in a seabird? PsyArXiv, 463hk, ver. 5 recommended and peer-reviewed by Peer Community In Ecology. doi: 10.31234/osf.io/463hk
[3] Morgan, C. L. (1894). An introduction to comparative psychology. London, UK: Walter Scott, Ltd. Retrieved from https://archive.org/details/introductiontoco00morg/page/53/mode/2up
[4] Meketa, I. (2014). A critique of the principle of cognitive simplicity in comparative cognition. Biology and Philosophy, 29(5), 731–745. doi: 10.1007/s10539-014-9429-z
[5] Fitzpatrick, S. (2017). Against Morgan's Canon. In K. Andrews and J. Beck (Eds.), The Routledge handbook of philosophy of animal minds (pp. 437–447). London, UK: Routledge, Taylor and Francis Group. doi: 10.4324/9781315742250.ch42
[6] Starzak, T. (2017). Interpretations without justification: a general argument against Morgan's Canon. Synthese, 194(5), 1681–1701. doi: 10.1007/s11229-016-1013-4
[7] Arbilly, M., and Lotem, A. (2017). Constructive anthropomorphism: a functional evolutionary approach to the study of human-like cognitive mechanisms in animals. Proceedings of the Royal Society B: Biological Sciences, 284(1865), 20171616. doi: 10.1098/rspb.2017.1616
[8] Taylor, A. H., Knaebe, B., and Gray, R. D. (2012). An end to insight? New Caledonian crows can spontaneously solve problems without planning their actions. Proceedings of the Royal Society B: Biological Sciences, 279(1749), 4977–4981. doi: 10.1098/rspb.2012.1998
[9] de Waal, F. (2016). Are we smart enough to know how smart animals are? New-York, USA: W. W. Norton and Company.
[10] Scheffer, M. (2014). The forgotten half of scientific thinking. Proceedings of the National Academy of Sciences, 111(17), 6119–6119. doi: 10.1073/pnas.1404649111
[11] Darwin, C. R. (1857). Letter to A. R. Wallace, 22 December 1857. Retrieved 30 January 2020, from https://www.darwinproject.ac.uk/letter/DCP-LETT-2192.xml
[12] Sol, D., Lefebvre, L., and Rodríguez-Teijeiro, J. D. (2005). Brain size, innovative propensity and migratory behaviour in temperate Palaearctic birds. Proceedings of the Royal Society B: Biological Sciences, 272(1571), 1433–1441. doi: 10.1098/rspb.2005.3099
| Evidence of tool use in a seabird? | Benjamin G. Farrar | Fayet, Hansen and Biro (1) provide two observations of Atlantic puffins, *Fratercula arctica*, performing self-directed actions while holding a stick in their beaks. The authors interpret this as evidence of tool use as they suggest that the stick... |  | Behaviour & Ethology | Francois-Xavier Dechaume-Moncharmont | | 2020-01-22 11:55:27 | View |
Explore and move: a key to success in a changing world?
Recommended by Blandine Doligez based on reviews by Joe Nocera, Marion Nicolaus and Laure Cauchard
Changes in the spatial range of many species are one of the major consequences of the profound alteration of environmental conditions due to human activities. Some species expand, sometimes spectacularly during invasions; others decline; some shift. Because these changes result in local biodiversity loss (whether local species go extinct or are replaced by colonizing ones), understanding the factors driving spatial range dynamics appears crucial to predict biodiversity dynamics. Identifying the factors that shape individual movement is a main step towards such understanding. The study described in this preregistration (McCune et al. 2020) falls within this context by testing possible links between individual exploration behaviour and movements related to daily space use in an avian study model currently rapidly expanding, the great-tailed grackle (Quiscalus mexicanus).
Movement and exploration: which direction(s) for the link between exploration and dispersal?
Individuals are known to differ in their tendency to explore the environment (Réale et al. 2007; Wolf and Weissing 2012) and therefore in their motivation to move. Accordingly, exploration has been shown to relate to dispersal behaviour, i.e. movements between breeding sites (Dingemanse et al. 2003, Le Galliard et al. 2011, Rasmussen and Belk 2012; reviews in Cote et al. 2010, Ronce et al. 2012). Yet, the mechanisms underlying this link often remain unclear, due to the correlative nature of the data. A classical assumption is that dispersers may benefit from a high capacity to explore, allowing them to familiarize quicker with their new environment once reached, thus alleviating dispersal costs (Bonte et al. 2012). The association between dispersal and exploration would in this case result from selection for this combination of traits (Ronce et al. 2012), even though dispersal event itself may be independent from (and precede the effect of) exploration behaviour. Alternatively (but not exclusively), dispersal may simply be the final outcome of longer movements by individuals exploring larger ranges (Badyaev et al. 1996, Schliehe-Diecks et al. 2012). In the absence of easy ways to manipulate dispersal behaviour, on the one hand, and exploration tendency, on the other hand, investigating detailed, small-scale individual movements in relation to exploration should thus shed light on which processes may yield the observed relations between exploration as an individual personality trait and large-scale, long-term movements, such as dispersal, underlying species range dynamics.
In this project, the exploration behaviour of grackles will be measured in controlled conditions using standardized tests in captivity (McCune et al. 2019) before individuals are released and their daily space use behaviour will then be measured using remote tracking over long time periods (McCune et al. 2020). Importantly, these coupled measures will be obtained for individuals captured in three different populations: within the historical range of the species, in the middle of its expanding range and at the edge of the range (McCune et al. 2020). Therefore, the project will test (i) whether daily space use of individuals is linked to their intrinsic exploration tendency and (ii) whether space use differs between individuals from different populations along the expanding range. The preregistration echoes a complementary project by the same team that will focus on exploration and test (iii) whether exploration tendency differs between individuals from these different populations. Taken together, these three analyses will therefore provide solid background information to assess the role of exploration in the individuals’ decisions leading to movement and range dynamics in this species.
As underlined in the preregistration, previous studies addressing the links between individual exploration behaviour and movements have mostly focused on dispersal. A first type of studies have (as will be done here) measured exploration behaviour of individuals, often in captivity (Dingemanse et al. 2003, Korsten et al. 2013) but also in the wild (Rasmussen and Belk 2012, Debeffe et al. 2013), and related these measures to subsequent dispersal behaviour. The (often implicit) underlying assumption is that more exploratory individuals will be more likely to move further, explore different habitats and thus end up breeding farther than less explorative ones. In other words, exploration tendency precedes and drives dispersal. Sometimes, exploratory behaviour is measured on individuals of known dispersal status, i.e. after the dispersal event (Hoset et al. 2011), in which case selection for certain exploration phenotypes among dispersers may already have occurred. Besides this first approach, another type of studies have measured ‘exploration’ behaviour under the form of prospecting movements of individuals and linked these movements to subsequent dispersal (often in the context of habitat selection). While these studies were in the past based on direct thus potentially biased observations (Reed et al. 1999), they now rely more and more on technological advances using (miniaturized) remote tracking devices (Ponchon et al. 2013) that provide far more complete and unbiased movement data, and sometimes also complementary measures of individuals’ internal state. In this case, the implicit assumption is that individuals prospecting farther and/or in more habitat patches will be more likely to settle in a site located farther away from their departure site, because of a more exhaustive sampling of possible sites allowing individuals to identify higher-quality sites (Badyaev et al. 1996). In other words, exploration tendency would not directly lead to higher movements or longer distances, but would allow individuals to optimize their habitat choice among more numerous options, thus leading to an increased dispersal probability or distance; the relation between exploration and dispersal would thus be indirect. Prospecting studies address more closely the underlying mechanisms of movement; however, they cannot easily separate intrinsic individual exploratory tendency from the prospecting movements themselves, with potential feedback effects of the information already gathered on future exploration of other sites or patches, thus on subsequent movements.
By focusing on individual daily space use movements as a mechanistic approach to understand large-scale movements potentially involved in colonization and range expansion, the grackle study described in this preregistration (McCune et al. 2020) will thus contribute to bridge the knowledge gaps between exploration and dispersal. By linking exploration measures obtained from a battery of standardized tests conducted in controlled conditions to individual daily space use and movements recorded in the wild, the grackle project is set in between previous studies addressing the links between exploration and dispersal: it will document exploration in a separate and independent context with respect to the movements themselves, and it will use a mechanistic view of detailed movements by the same individuals in the wild to explore potential implications for dispersal and range expansion. Testing differences between the three study populations over the species range will indeed inform about potential large-scale, population implications of among-individual variation in the link between exploration and movements. Because this study will only measure already settled adult individuals whose previous history is unknown, there will nevertheless be no direct possible exploration of the link with either previous or subsequent dispersal behaviour. Thus, the potential links studied here relate more directly to post-dispersal benefits of exploration for an optimal exploitation of the new environment. Yet, if exploration is a life-long personality trait linked to daily movement patterns, it may also relate to natal dispersal movements in young individuals.
Evolutionary and conservation perspectives
If the results of the project reveal that exploration tendency and daily space use movements are indeed linked, and that individuals from populations across the species range differ in these traits, new questions will emerge. A first question would be whether such among-individual differences are at the origin of range expansion or rather one of its consequences since, again, we deal with correlative data here. In other words, individuals may differ in exploration tendency, and this may confer them different ability to move around, find and colonize new habitats; or individuals may show differences in exploration following arrival in a new habitat, either because more explorative individuals gain fitness benefits and are thus selected, or because of behavioural plasticity and post-colonization adjustment of exploration behaviour when facing new ecological and social conditions in the new environment. Another open question relates to the link between daily space use and dispersal: is dispersal a by-product of higher daily movements that allow individuals to discover new favorable places where to settle? Exploring this link could involve measuring just fledged individuals before natal dispersal occurs and/or individuals chosen according to their own dispersal history, and this would then imply long-term population monitoring as an efficient (but constraining) tool to address such questions. Finally, assessing the fitness consequences of the link between exploration and space use behaviour, and whether these consequences differ between populations along the range expansion, would also be needed to understand the contribution of this link to the invasion success of this species.
The study model chosen for this project is a rapidly expanding species. Importantly, however, and as emphasized in the preregistration, documenting links between exploration and daily space use patterns as well as differences between populations with different trajectories can provide crucial information in general to understand population persistence in response to global climate and landscape changes, both regarding invasion ability or extinction risk. The information should be key to assess the probability that a species may decline, persist or expand in studies addressing biodiversity and community dynamics in a changing world.
References
Badayev, A. V., Martin, T. E and Etges, W. J. 1996. Habitat sampling and habitat selection by female wild turkeys: ecological correlates and reproductive consequences. Auk 113: 636-646. doi: https://doi.org/10.2307/4088984
Bonte, D. et al. 2012. Costs of dispersal. Biological Reviews 87: 290-312. doi: https://doi.org/10.1111/j.1469-185X.2011.00201.x
Cote, J., Clobert, J., Brodin, T., Fogarty, S. and Sih, A. 2010. Personality-dependent dispersal: characterization, ontogeny and consequences for spatially structured populations. Philosophical Transactions of the Royal Society B 365: 4065-4576. doi: https://doi.org/10.1098/rstb.2010.0176
Debeffe, L., Morellet, N., Cargnelutti, B., Lourtet, B., Coulon, A., Gaillard, J.-M., Bon, R. and Hewison A. J. M. 2013. Exploration as a key component of natal dispersal: dispersers explore more than philopatric individuals in roe deer. Animal Behaviour 86: 143-151. doi: https://doi.org/10.1016/j.anbehav.2013.05.005
Dingemanse, N. J., Both, C., van Noordwijk, A. J., Rutten, A. L. and Drent, P. J. 2003. Natal dispersal and personalities in great tits (Parus major). Proceedings of the Royal Society B 270: 741-747. doi: https://doi.org/10.1098/rspb.2002.2300
Hoset, K. S., Ferchaud, A.-L., Dufour, F., Mersch, D., Cote, J. and Le Galliard, J.-F. 2011. Natal dispersal correlates with behavioral traits that are not consistent across early life stages. Behavioral Ecology 22: 176–183. doi: https://doi.org/10.1093/beheco/arq188
Korsten, P., van Overveld, T., Adriaensen, F. and Matthysen, E. 2013. Genetic integration of local dispersal and exploratory behaviour in a wild bird. Nature Communications 4: 2362. doi: https://doi.org/10.1038/ncomms3362
Le Galliard, J.-F., Rémy, A., Ims, R. A. and Lambin, X. 2011. Patterns and processes of dispersal behaviour in arvicoline rodents. Molecular Ecology 21: 505-523. doi: https://doi.org/10.1111/j.1365-294X.2011.05410.x
McCune K, Ross C, Folsom M, Bergeron L, Logan CJ. 2020. Does space use behavior relate to exploration in a species that is rapidly expanding its geographic range? http://corinalogan.com/Preregistrations/gspaceuse.html In principle acceptance by PCI Ecology of the version on 23 Sep 2020 https://github.com/corinalogan/grackles/blob/master/Files/Preregistrations/gspaceuse.Rmd.
McCune K, MacPherson M, Rowney C, Bergeron L, Folsom M, Logan CJ. 2019. Is behavioral flexibility linked with exploration, but not boldness, persistence, or motor diversity? (http://corinalogan.com/Preregistrations/gexploration.html) In principle acceptance by PCI Ecology of the version on 27 Mar 2019 https://github.com/corinalogan/grackles/blob/master/Files/Preregistrations/gexploration.Rmd
Ponchon, A., Grémillet, D., Doligez, B., Chambert, T., Tveraa, T., González-Solís, J. and Boulinier, T. 2013. Tracking prospecting movements involved in breeding habitat selection: insights, pitfalls and perspectives. Methods in Ecology and Evolution 4: 143-150. doi: https://doi.org/10.1111/j.2041-210x.2012.00259.x
Rasmussen, J. E. and Belk, M. C. 2012. Dispersal behavior correlates with personality of a North American fish. Current Zoology 58: 260–270. doi: https://doi.org/10.1093/CZOOLO%2F58.2.260
Réale, D., Reader, S. M., Sol, D., McDougall, P. T. and Dingemanse, N. J. 2007. Integrating animal temperament within ecology and evolution. Biological Reviews 82: 291-318. doi: https://doi.org/10.1111/j.1469-185x.2007.00010.x
Reed, J. M., Boulinier, T., Danchin, E. and Oring, L. W. 1999. Informed dispersal: prospecting by birds for breeding sites. Current Ornithology 15: 189-259. doi: https://doi.org/10.1007/978-1-4757-4901-4_5
Ronce, O. and Clobert, J. 2012. Dispersal syndromes. pp. 119-138 In Dispersal Ecology and Evolution (eds. Clobert, J., Baguette, M., Benton, T. G. and Bullock, J. M.), pp. 119-138. Oxford University Press.
Schliehe-Diecks, S., Eberle, M. and Kappeler, P. M. 2012. Walk the line - dispersal movements of gray mouse lemurs (Microcebus murinus). Behavioral Ecology and Sociobiology 66: 1175-1185. doi: https://dx.doi.org/10.1007%2Fs00265-012-1371-y
Wolf, M. and Weissing, F. J. 2012. Animal personalities: consequences for ecology and evolution. Trends in Ecology and Evolution 27: 452-461. doi: https://doi.org/10.1016/j.tree.2012.05.001
| Does space use behavior relate to exploration in a species that is rapidly expanding its geographic range? | Kelsey B. McCune, Cody Ross, Melissa Folsom, Luisa Bergeron, Corina Logan | Great-tailed grackles (Quiscalus mexicanus) are rapidly expanding their geographic range (Wehtje 2003). Range expansion could be facilitated by consistent behavioural differences between individuals on the range edge and those in other parts of th... |  | Behaviour & Ethology, Biological invasions, Conservation biology, Habitat selection, Phenotypic plasticity, Preregistrations, Spatial ecology, Metacommunities & Metapopulations | Blandine Doligez | | 2019-09-30 19:27:40 | View |
A community perspective on the concept of marine holobionts: current status, challenges, and future directions
Simon M. Dittami, Enrique Arboleda, Jean-Christophe Auguet, Arite Bigalke, Enora Briand, Paco Cárdenas, Ulisse Cardini, Johan Decelle, Aschwin Engelen, Damien Eveillard, Claire M.M. Gachon, Sarah Griffiths, Tilmann Harder, Ehsan Kayal, Elena Kazamia, Francois H. Lallier, Mónica Medina, Ezequiel M. Marzinelli, Teresa Morganti, Laura Núñez Pons, Soizic Pardo, José Pintado Valverde, Mahasweta Saha, Marc-André Selosse, Derek Skillings, Willem Stock, Shinichi Sunagawa, Eve Toulza, Alexey Vorobev, Cat...
10.5281/zenodo.3696771
Marine holobiont in the high throughput sequencing era
Recommended by Sophie Arnaud-Haond and Corinne Vacher based on reviews by Sophie Arnaud-Haond and Aurélie Tasiemski
The concept of holobiont dates back to more than thirty years, it was primarily coined to hypothesize the importance of symbiotic associations to generate significant evolutionary novelties. Quickly adopted to describe the now well-studied system formed by zooxanthella associated corals, this concept expanded much further after the emergence of High-Throughput Sequencing and associated progresses in metabarcoding and metagenomics.
Holobionts – defined as the association between an individual host and its microbiota - are now increasingly described at sea and on land. The opinion article by Dittami et al. [1] provides a synthetic overview of marine holobionts. It retraces the history of the holobiont concept, recalls the main mechanisms underlying the association between hosts and microbial communities, highlights the influence of these symbioses on marine ecosystem functioning, and outlines current tools and future lines of research.
In particular, the article discusses some particularities of marine systems, such as the strong connectivity allowing an exchange of microorganisms and chemical signals between and within holobionts.
The authors advocate the need to bridge the gap between large scale exploration studies and smaller scale mechanistic studies, by conducting interdisciplinary research (combining physiology, biochemistry, ecology, experimentation and computational modeling) on some keystone holobionts.
Finally, one strength of the paper by Dittami et al. [1] is that it places the concept of the holobiont in an applied research framework. Several possible applications of knowledge on host-microbiota interactions are suggested, both in the field of aquaculture and that of monitoring the health of marine ecosystems. This article contains all the necessary elements for someone who would like to jump into the study of the holobionths in the marine world.
References
[1] Dittami SM, Arboleda E, Auguet J, Bigalke A, Briand E, Cardenas P, Cardini U, Decelle J, Engelen AH, Eveillard D, Gachon CMM, Griffiths SM, Harder T, Kayal E, Kazamia E, Lallier FH, Medina M, Marzinelli E, Morganti T, Núñez Pons L, Prado S, Pintado J, Saha M, Selosse M, Skillings D, Stock W, Sunagawa S, Toulza E, Vorobev A, Leblanc C, Not F. (2020). A community perspective on the concept of marine holobionts: current status, challenges, and future directions. Zenodo, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.5281/zenodo.3696771
| A community perspective on the concept of marine holobionts: current status, challenges, and future directions | Simon M. Dittami, Enrique Arboleda, Jean-Christophe Auguet, Arite Bigalke, Enora Briand, Paco Cárdenas, Ulisse Cardini, Johan Decelle, Aschwin Engelen, Damien Eveillard, Claire M.M. Gachon, Sarah Griffiths, Tilmann Harder, Ehsan Kayal, Elena Kazam... | Host-microbe interactions play crucial roles in marine ecosystems. However, we still have very little understanding of the mechanisms that govern these relationships, the evolutionary processes that shape them, and their ecological consequences. T... |  | Marine ecology, Microbial ecology & microbiology, Symbiosis | Sophie Arnaud-Haond | | 2019-02-05 17:57:11 | View |
Does elevated parasite richness in the environment affect daily path length of animals or is it the converse? An answer bringing some new elements of discussion
Recommended by Cédric Sueur based on reviews by 2 anonymous reviewers
In 2015, Brockmeyer et al. [1] suggested that mandrills (Mandrillus sphinx) may accept additional ranging costs to avoid heavily parasitized areas. Following this paper, Bicca-Marques and Calegaro-Marques [2] questioned this interpretation and presented other hypotheses. To summarize, whilst Brockmeyer et al. [1] proposed that elevated daily path length may be a consequence of elevated parasite richness, Bicca-Marques and Calegaro-Marques [2] viewed it as a cause. In this current paper, Charpentier and Kappeler [3] respond to some of the criticisms by Bicca-Marques and Calegaro-Marques and discuss the putative parsimony of the two competing scenarios. The manuscript is interesting and focuses on an important question concerning the discussion about the social organization and home range use in wild mandrills. This answer helps to move this debate forward and should stimulate more empirical studies of the role of environmentally-transmitted parasites in shaping ranging and movement patterns of wild vertebrates. Given the elements this paper brings to the topics, it should have been published in American Journal of Primatology, the journal that published the two previous articles.
References
[1] Brockmeyer, T., Kappeler, P. M., Willaume, E., Benoit, L., Mboumba, S., & Charpentier, M. J. E. (2015). Social organization and space use of a wild mandrill (Mandrillus sphinx) group. American Journal of Primatology, 77(10), 1036–1048. doi: 10.1002/ajp.22439
[2] Bicca-Marques, J. C., & Calegaro-Marques, C. (2016). Ranging behavior drives parasite richness: A more parsimonious hypothesis. American Journal of Primatology, 78(9), 923–927. doi: 10.1002/ajp.22561
[3] Charpentier, M. J., & Kappeler, P. M. (2018). A reply to “Ranging Behavior Drives Parasite Richness: A More Parsimonious Hypothesis.” ArXiv:1805.08151v2 [q-Bio]. Retrieved from http://arxiv.org/abs/1805.08151
| A reply to “Ranging Behavior Drives Parasite Richness: A More Parsimonious Hypothesis” | Charpentier MJE, Kappeler PM | In a recent article, Bicca-Marques and Calegaro-Marques [2016] discussed the putative assumptions related to an interpretation we provided regarding an observed positive relationship between weekly averaged parasite richness of a group of mandrill... |  | Behaviour & Ethology, Evolutionary ecology, Foraging, Host-parasite interactions, Spatial ecology, Metacommunities & Metapopulations, Zoology | Cédric Sueur | | 2018-05-22 10:59:33 | View |
Investigating sex differences in genetic relatedness in great-tailed grackles in Tempe, Arizona to infer potential sex biases in dispersal
August Sevchik, Corina Logan, Melissa Folsom, Luisa Bergeron, Aaron Blackwell, Carolyn Rowney, Dieter Lukas
http://corinalogan.com/Preregistrations/gdispersal.html
Investigate fine scale sex dispersal with spatial and genetic analyses
Recommended by Sophie Beltran-Bech based on reviews by Sylvine Durand and 1 anonymous reviewer
The preregistration "Investigating sex differences in genetic relatedness in great-tailed grackles in Tempe, Arizona to infer potential sex biases in dispersal" [1] presents the analysis plan that will be used to genetically and spatially investigate sex-biased dispersal in great-tailed grackles (Quiscalus mexicanus).
Several hypotheses implying mating systems, intrasexual competition or sex-related handicaps have been proposed to explain the diversity of dispersal patterns between or within species according to their ecological requirements, environmental factors such as seasonality [2], or individual characteristics such as age [3] or sex [4].
In birds, females are classically the dispersing sex, while males remain close to the place they were hatched [5], with potential benefits that males derive from knowing the local environment to establish territories [6].
In great-tailed grackles the males hold territories and the females choose which territory to place their nest in [7]. In this context, the main hypothesis is that females are the dispersing sex in this species. The authors of this preregistration plan to investigate this hypothesis and its 3 alternatives ((i) the males are the dispersing sex, (ii) both sexes disperse or (iii) neither of the two sexes disperse), investigating the spatial distribution of genetic relatives.
The authors plan to measure the genetic relatedness (using SNP markers) and geographic distances among all female dyads and among all male dyads in the fine geographic scale (Tempe campus, Arizona). If females disperse away from relatives, the females will be less likely to be found geographically close to genetic relatives.
This pre-registration shows that the authors are well aware of the possible limitations of their study, particularly in relation to their population of 57 individuals, on a small scale. But they will use methods that should be able to detect a signal. They were very good at incorporating the reviewers' comments and suggestions, which enabled them to produce a satisfactory and interesting version of the manuscript presenting their hypotheses, limitations and the methods they plan to use. Another point I would like to stress is that this pre-registration practice is a very good one that makes it possible to anticipate the challenges and the type of analyses to be carried out, in particular by setting out the working hypotheses and confronting them (as well as the methods envisaged) with peers from this stage. I therefore recommend this manuscript and thank all the contributors (authors and reviewers) for their work. I look forward to seeing the outcomes of this study.
References
[1] Sevchik A., Logan C. J., Folsom M., Bergeron L., Blackwell A., Rowney C., and Lukas D. (2019). Investigating sex differences in genetic relatedness in great-tailed grackles in Tempe, Arizona to infer potential sex biases in dispersal. In principle recommendation by Peer Community In Ecology. corinalogan.com/Preregistrations/gdispersal.html
[2] Fies, M. L., Puckett, K. M., and Larson-Brogdon, B. (2002). Breeding season movements and dispersal of Northern Bobwhites in fragmented habitats of Virginia. Vol. 5 , Article 35. Available at: trace.tennessee.edu/nqsp/vol5/iss1/35
[3] Marvá, M., and San Segundo, F. (2018). Age-structure density-dependent fertility and individuals dispersal in a population model. Mathematical biosciences, 300, 157-167. doi: 10.1016/j.mbs.2018.03.029
[4] Trochet, A., Courtois, E. A., Stevens, V. M., Baguette, M., Chaine, A., Schmeller, D. S., Clobert, J., and Wiens, J. J. (2016). Evolution of sex-biased dispersal. The Quarterly Review of Biology, 91(3), 297-320. doi: 10.1086/688097
[5] Greenwood, P. J., and Harvey, P. H. (1982). The natal and breeding dispersal of birds. Annual review of ecology and systematics, 13(1), 1-21. doi: 10.1146/annurev.es.13.110182.000245
[6] Greenwood, P. J. (1980). Mating systems, philopatry and dispersal in birds and mammals. Animal behaviour, 28(4), 1140-1162. doi: 10.1016/S0003-3472(80)80103-5
[7] Johnson, K., DuVal, E., Kielt, M., and Hughes, C. (2000). Male mating strategies and the mating system of great-tailed grackles. Behavioral Ecology, 11(2), 132-141. doi: 10.1093/beheco/11.2.132
| Investigating sex differences in genetic relatedness in great-tailed grackles in Tempe, Arizona to infer potential sex biases in dispersal | August Sevchik, Corina Logan, Melissa Folsom, Luisa Bergeron, Aaron Blackwell, Carolyn Rowney, Dieter Lukas | In most bird species, females disperse prior to their first breeding attempt, while males remain close to the place they were hatched for their entire lives (Greenwood and Harvey (1982)). Explanations for such female bias in natal dispersal have f... |  | Behaviour & Ethology, Life history, Preregistrations, Social structure, Zoology | Sophie Beltran-Bech | | 2019-07-24 12:47:07 | View |
Diapause is not selected as a bet-hedging strategy in insects: a meta-analysis of reaction norm shapes
Jens Joschinski and Dries Bonte
10.1101/752881
When to diapause or not to diapause? Winter predictability is not the answer
Recommended by Bastien Castagneyrol based on reviews by Kévin Tougeron, Md Habibur Rahman Salman and 1 anonymous reviewer
Winter is a harsh season for many organisms that have to cope with food shortage and potentially lethal temperatures. Many species have evolved avoidance strategies. Among them, diapause is a resistance stage many insects use to overwinter. For an insect, it is critical to avoid lethal winter temperatures and thus to initiate diapause before winter comes, while making the most of autumn suitable climatic conditions [1,2]. Several cues can be used to appreciate that winter is coming, including day length and temperature [3]. But climate changes, temperatures rise and become more variable from year to year, which imposes strong pressure upon insect phenology [4]. How can insects adapt to changes in the mean and variance of winter onset?
In this paper, Jens Joschinski and Dries Bonte [5] address this question by using a well conducted meta-analysis of 458 diapause reaction norms obtained from 60 primary studies. They first ask first if insect mean diapause timing is tuned to match winter onset. They further ask if insects adapt to climatic unpredictability through a bet-hedging strategy by playing it safe and avoid risk (conservative bet-hedging) or on the contrary by avoiding to put all their eggs in one basket and spread the risk among their offspring (diversified bet-hedging). From published papers, the authors extracted data on mean diapause timing and information on latitude from which they retrieved day length inducing diapause, the date of winter onset and the day length at winter onset.
They found a positive correlation between latitude and the day length inducing diapause. On the contrary they found positive but (very) weak correlation between the date of winter onset and the date of diapause, thus indicating that diapause timing is not as optimally adapted to local environments as expected, particularly at high latitudes. They only found weak correlations between climate unpredictability and variability in diapause timing, and no correlation between climate unpredictability and deviation from optimal diapause timing. Together, these findings go against the hypothesis that insects use diversified or conservative bet-hedging strategies to cope with uncertainty in climatic conditions.
This is what makes the study thought provoking: the results do not match the theory well. Not because of a lack of data or a narrow scope, but because diapause is a complex trait that is determined by a large array of physiological and ecological factors [3]. Determining what are these factors is of particular interest in the face of the current climate change. This study shows what does not determine the timing of insect diapause. Researchers now know where to look at to improve our understanding of this key aspect of insect adaptation to climatic conditions.
References
[1] Dyck, H. V., Bonte, D., Puls, R., Gotthard, K., and Maes, D. (2015). The lost generation hypothesis: could climate change drive ectotherms into a developmental trap? Oikos, 124(1), 54–61. doi: 10.1111/oik.02066
[2] Gallinat, A. S., Primack, R. B., and Wagner, D. L. (2015). Autumn, the neglected season in climate change research. Trends in Ecology & Evolution, 30(3), 169–176. doi: 10.1016/j.tree.2015.01.004
[3] Tougeron, K. (2019). Diapause research in insects: historical review and recent work perspectives. Entomologia Experimentalis et Applicata, 167(1), 27–36. doi: 10.1111/eea.12753
[4] Bale, J. S., and Hayward, S. a. L. (2010). Insect overwintering in a changing climate. Journal of Experimental Biology, 213(6), 980–994. doi: 10.1242/jeb.037911
[5] Joschinski, J., and Bonte, D. (2020). Diapause is not selected as a bet-hedging strategy in insects: a meta-analysis of reaction norm shapes. BioRxiv, 752881, ver. 3 recommended and peer-reviewed by PCI Ecology. doi: 10.1101/752881
| Diapause is not selected as a bet-hedging strategy in insects: a meta-analysis of reaction norm shapes | Jens Joschinski and Dries Bonte | Many organisms escape from lethal climatological conditions by entering a resistant resting stage called diapause, and it is essential that this strategy remains optimally timed with seasonal change. Climate change therefore exerts selection press... | 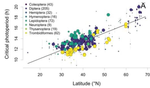 | Maternal effects, Meta-analyses, Phenotypic plasticity, Terrestrial ecology | Bastien Castagneyrol | | 2019-09-20 11:47:47 | View |
Are condition indices positively related to each other and to fitness?: a test with grackles
Recommended by Marcos Mendez based on reviews by Javier Seoane and Isabel López-Rull
Reproductive succes, as a surrogate of individual fitness, depends both on extrinsic and intrinsic factors [1]. Among the intrinsic factors, resource level or health are considered important potential drivers of fitness but exceedingly difficult to measure directly. Thus, a host of proxies have been suggested, known as condition indices [2]. The question arises whether all condition indices consistently measure the same "inner state" of individuals and whether all of them similarly correlate to individual fitness. In this preregistration, Berens and colleagues aim to answer this question for two common condition indices, fat score and scaled mass index (Fig. 1), using great-tailed grackles as a model system. Although this question is not new, it has not been satisfactorily solved and both reviewers found merit in the attempt to clarify this matter.

Figure 1. Hypothesized relationships between two condition indices and reproductive success. Single arrow heads indicate causal relationships; double arrow heads indicate only correlation. In a best case scenario, all relationships should be positive and linear.
A problem in adressing this question with grackles is limited population, ergo sample, size and limited possibilites of recapture individuals. Some relationships can be missed due to low statistical power. Unfortunately, existing tools for power analysis fall behind complex designs and the one planned for this study. Thus, any potentially non significant relationship has to be taken cautiously. Nevertheless, even if grackles will not provide a definitive answer (they never meant to do it), this preregistration can inspire broader explorations of matches and mismatches across condition indices and species, as well as uncover non-linear relationships with reproductive success.
References
[1] Roff, D. A. (2001). Life history evolution. Oxford University Press, Oxford.
[2] Labocha, M. K.; Hayes, J. P. (2012). Morphometric indices of body condition in birds: a review. Journal of Ornithology 153: 1–22. doi: 10.1007/s10336-011-0706-1
| Validating morphological condition indices and their relationship with reproductive success in great-tailed grackles | Jennifer M. Berens, Corina J. Logan, Melissa Folsom, Luisa Bergeron, Kelsey B. McCune | Morphological variation among individuals has the potential to influence multiple life history characteristics such as dispersal, migration, reproductive fitness, and survival (Wilder, Raubenheimer, and Simpson (2016)). Theoretically, individuals ... |  | Behaviour & Ethology, Conservation biology, Demography, Morphometrics, Preregistrations, Zoology | Marcos Mendez | | 2019-08-05 20:05:56 | View |
Riparian forest restoration as sources of biodiversity and ecosystem functions in anthropogenic landscapes
Yasmine Antonini, Marina Vale Beirao, Fernanda Vieira Costa, Cristiano Schetini Azevedo, Maria Wojakowski, Alessandra Kozovits, Maria Rita Silverio Pires, Hildeberto Caldas Sousa, Maria Cristina Teixeira Braga Messias, Maria Augusta Goncalves Fujaco, Mariangela Garcia Praca Leite, Joice Paiva Vidigal Martins, Graziella Franca Monteiro, Rodolfo Dirzo
https://doi.org/10.1101/2021.09.08.459375
Complex but positive diversity - ecosystem functioning relationships in Riparian tropical forests
Recommended by Werner Ulrich based on reviews by 2 anonymous reviewers
Many ecological drivers can impact ecosystem functionality and multifunctionality, with the latter describing the joint impact of different functions on ecosystem performance and services. It is now generally accepted that taxonomically richer ecosystems are better able to sustain high aggregate functionality measures, like energy transfer, productivity or carbon storage (Buzhdygan 2020, Naeem et al. 2009), and different ecosystem services (Marselle et al. 2021) than those that are less rich. Antonini et al. (2022) analysed an impressive dataset on animal and plant richness of tropical riparian forests and abundances, together with data on key soil parameters. Their work highlights the importance of biodiversity on functioning, while accounting for a manifold of potentially covarying drivers. Although the key result might not come as a surprise, it is a useful contribution to the diversity - ecosystem functioning topic, because it is underpinned with data from tropical habitats. To date, most analyses have focused on temperate habitats, using data often obtained from controlled experiments.
The paper also highlights that diversity–functioning relationships are complicated. Drivers of functionality vary from site to site and each measure of functioning, including parameters as demonstrated here, can be influenced by very different sets of predictors, often associated with taxonomic and trait diversity. Single correlative comparisons of certain aspects of diversity and functionality might therefore return very different results. Antonini et al. (2022) show that, in general, using 22 predictors of functional diversity, varying predictor subsets were positively associated with soil functioning. Correlational analyses alone cannot resolve the question of causal link. Future studies should therefore focus on inferring precise mechanisms behind the observed relationships, and the environmental constraints on predictor subset composition and strength.
References
Antonini Y, Beirão MV, Costa FV, Azevedo CS, Wojakowski MM, Kozovits AR, Pires MRS, Sousa HC de, Messias MCTB, Fujaco MA, Leite MGP, Vidigal JP, Monteiro GF, Dirzo R (2022) Riparian forest restoration as sources of biodiversity and ecosystem functions in anthropogenic landscapes. bioRxiv, 2021.09.08.459375, ver. 3 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2021.09.08.459375
Buzhdygan OY, Meyer ST, Weisser WW, Eisenhauer N, Ebeling A, Borrett SR, Buchmann N, Cortois R, De Deyn GB, de Kroon H, Gleixner G, Hertzog LR, Hines J, Lange M, Mommer L, Ravenek J, Scherber C, Scherer-Lorenzen M, Scheu S, Schmid B, Steinauer K, Strecker T, Tietjen B, Vogel A, Weigelt A, Petermann JS (2020) Biodiversity increases multitrophic energy use efficiency, flow and storage in grasslands. Nature Ecology & Evolution, 4, 393–405. https://doi.org/10.1038/s41559-020-1123-8
Marselle MR, Hartig T, Cox DTC, de Bell S, Knapp S, Lindley S, Triguero-Mas M, Böhning-Gaese K, Braubach M, Cook PA, de Vries S, Heintz-Buschart A, Hofmann M, Irvine KN, Kabisch N, Kolek F, Kraemer R, Markevych I, Martens D, Müller R, Nieuwenhuijsen M, Potts JM, Stadler J, Walton S, Warber SL, Bonn A (2021) Pathways linking biodiversity to human health: A conceptual framework. Environment International, 150, 106420. https://doi.org/10.1016/j.envint.2021.106420
Naeem S, Bunker DE, Hector A, Loreau M, Perrings C (Eds.) (2009) Biodiversity, Ecosystem Functioning, and Human Wellbeing: An Ecological and Economic Perspective. Oxford University Press, Oxford. https://doi.org/10.1093/acprof:oso/9780199547951.001.0001
| Riparian forest restoration as sources of biodiversity and ecosystem functions in anthropogenic landscapes | Yasmine Antonini, Marina Vale Beirao, Fernanda Vieira Costa, Cristiano Schetini Azevedo, Maria Wojakowski, Alessandra Kozovits, Maria Rita Silverio Pires, Hildeberto Caldas Sousa, Maria Cristina Teixeira Braga Messias, Maria Augusta Goncalves Fuja... | <ol>
<li style="text-align: justify;">Restoration of tropical riparian forests is challenging, since these ecosystems are the most diverse, dynamic, and complex physical and biological terrestrial habitats. This study tested whether biodiversity ... |  | Biodiversity, Community ecology, Ecological successions, Ecosystem functioning, Terrestrial ecology | Werner Ulrich | | 2021-09-10 10:51:23 | View |
Crown defoliation decreases reproduction and wood growth in a marginal European beech population.
Sylvie Oddou-Muratorio, Cathleen Petit-Cailleux, Valentin Journé, Matthieu Lingrand, Jean-André Magdalou, Christophe Hurson, Joseph Garrigue, Hendrik Davi, Elodie Magnanou.
https://doi.org/10.1101/474874
Defoliation induces a trade-off between reproduction and growth in a southern population of Beech
Recommended by Georges Kunstler based on reviews by 3 anonymous reviewers
Individuals ability to withstand abiotic and biotic stresses is crucial to the maintenance of populations at climate edge of tree species distribution. We start to have a detailed understanding of tree growth response and to a lesser extent mortality response in these populations. In contrast, our understanding of the response of tree fecundity and recruitment remains limited because of the difficulty to monitor it at the individual tree level in the field. Tree recruitment limitation is, however, crucial for tree population dynamics [1-2].
In their study Oddou-Muratorio et al. [3] use a new method that they recently developed that jointly estimate male and female effective fecundity in natural populations using naturally established seedlings [4]. Their method uses a spatially explicit Bayesian analysis based on molecular markers and parentage analyses (MEMM program [4]). They apply this method to an unmanaged Beech forest at the southern edge of Beech distribution, where tree defoliation – taken as an integrative indicator of tree abiotic and biotic stress – and growth have been monitored for all adult trees.
This allows the authors to explore alternative hypothesis about tree fecundity response to stress. In one hand, biotic and abiotic stresses are thought to negatively impact tree fecundity. In the other hand, management and studies of orchard fruit tree support the idea that stress could trigger a compensatory increase of fecundity at the cost of other performances such as growth and survival.
They show that both growth and female fecundity are negatively affected by defoliation. There was no evidence that stresses trigger a compensatory increase of fecundity. Yet, they also found that, for large highly defoliated trees, there was a trade-off between growth and female fecundity. Some individuals are able to mitigate stress impact on fecundity by decreasing their growth. It is difficult to understand with available data what is driving such divergent responses between defoliated individuals. This could be related to differences in micro-environmental conditions or genetic background of individual trees. Such individual-level difference in response to stress could be crucial to understand tree populations response to ongoing climate change. This study clearly opens exciting new perspectives to apply such new methods to understand the role of fecundity on tree population dynamics. For instance, could we apply this method across the species distribution to understand how effective fecundity and its response to abiotic stress change between southern edge populations, core populations, and northern edge populations? Using time-series retrieved from such analysis can we disentangle the effect of different climatic drivers? It would also be interesting to see how such results can contribute to analyses covering the full tree life cycle to understand the contribution of fecundity response to population and evolutionary.
References
[1] Clark, J. S. et al. (1999). Interpreting recruitment limitation in forests. American Journal of Botany, 86(1), 1-16. doi: 10.2307/2656950
[2] Petit, R. J., and Hampe, A. (2006). Some evolutionary consequences of being a tree. Annu. Rev. Ecol. Evol. Syst., 37, 187-214. doi: 10.1146/annurev.ecolsys.37.091305.110215
[3] Oddou-Muratorio, S., Petit, C., Journe, V., Lingrand, M., Magdalou, J. A., Hurson, C., Garrigue, J., Davi, H. and Magnanou, E. (2019). Crown defoliation decreases reproduction and wood growth in a marginal European beech population. bioRxiv, 474874, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/474874
[4] Oddou‐Muratorio, S. and Klein, E. K. (2008). Comparing direct vs. indirect estimates of gene flow within a population of a scattered tree species. Molecular Ecology, 17(11), 2743-2754. doi: 10.1111/j.1365-294X.2008.03783.x
| Crown defoliation decreases reproduction and wood growth in a marginal European beech population. | Sylvie Oddou-Muratorio, Cathleen Petit-Cailleux, Valentin Journé, Matthieu Lingrand, Jean-André Magdalou, Christophe Hurson, Joseph Garrigue, Hendrik Davi, Elodie Magnanou. | <p>1. Although droughts and heatwaves have been associated to increased crown defoliation, decreased growth and a higher risk of mortality in many forest tree species, their impact on tree reproduction and forest regeneration still remains underst... |  | Climate change, Eco-evolutionary dynamics, Molecular ecology, Physiology, Population ecology | Georges Kunstler | | 2018-11-20 13:29:42 | View |







