
KUNSTLER Georges
- LESSEM, Irstea, St-Martin-d’Hères, France
- Biodiversity, Biogeography, Climate change, Coexistence, Community ecology, Competition, Demography, Ecological successions, Facilitation & Mutualism, Macroecology, Population ecology, Terrestrial ecology, Theoretical ecology
- recommender
Recommendation: 1
Review: 1
Recommendation: 1
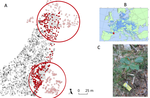
Crown defoliation decreases reproduction and wood growth in a marginal European beech population.
Defoliation induces a trade-off between reproduction and growth in a southern population of Beech
Recommended by Georges Kunstler based on reviews by 3 anonymous reviewersIndividuals ability to withstand abiotic and biotic stresses is crucial to the maintenance of populations at climate edge of tree species distribution. We start to have a detailed understanding of tree growth response and to a lesser extent mortality response in these populations. In contrast, our understanding of the response of tree fecundity and recruitment remains limited because of the difficulty to monitor it at the individual tree level in the field. Tree recruitment limitation is, however, crucial for tree population dynamics [1-2].
In their study Oddou-Muratorio et al. [3] use a new method that they recently developed that jointly estimate male and female effective fecundity in natural populations using naturally established seedlings [4]. Their method uses a spatially explicit Bayesian analysis based on molecular markers and parentage analyses (MEMM program [4]). They apply this method to an unmanaged Beech forest at the southern edge of Beech distribution, where tree defoliation – taken as an integrative indicator of tree abiotic and biotic stress – and growth have been monitored for all adult trees.
This allows the authors to explore alternative hypothesis about tree fecundity response to stress. In one hand, biotic and abiotic stresses are thought to negatively impact tree fecundity. In the other hand, management and studies of orchard fruit tree support the idea that stress could trigger a compensatory increase of fecundity at the cost of other performances such as growth and survival.
They show that both growth and female fecundity are negatively affected by defoliation. There was no evidence that stresses trigger a compensatory increase of fecundity. Yet, they also found that, for large highly defoliated trees, there was a trade-off between growth and female fecundity. Some individuals are able to mitigate stress impact on fecundity by decreasing their growth. It is difficult to understand with available data what is driving such divergent responses between defoliated individuals. This could be related to differences in micro-environmental conditions or genetic background of individual trees. Such individual-level difference in response to stress could be crucial to understand tree populations response to ongoing climate change. This study clearly opens exciting new perspectives to apply such new methods to understand the role of fecundity on tree population dynamics. For instance, could we apply this method across the species distribution to understand how effective fecundity and its response to abiotic stress change between southern edge populations, core populations, and northern edge populations? Using time-series retrieved from such analysis can we disentangle the effect of different climatic drivers? It would also be interesting to see how such results can contribute to analyses covering the full tree life cycle to understand the contribution of fecundity response to population and evolutionary.
References
[1] Clark, J. S. et al. (1999). Interpreting recruitment limitation in forests. American Journal of Botany, 86(1), 1-16. doi: 10.2307/2656950
[2] Petit, R. J., and Hampe, A. (2006). Some evolutionary consequences of being a tree. Annu. Rev. Ecol. Evol. Syst., 37, 187-214. doi: 10.1146/annurev.ecolsys.37.091305.110215
[3] Oddou-Muratorio, S., Petit, C., Journe, V., Lingrand, M., Magdalou, J. A., Hurson, C., Garrigue, J., Davi, H. and Magnanou, E. (2019). Crown defoliation decreases reproduction and wood growth in a marginal European beech population. bioRxiv, 474874, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/474874
[4] Oddou‐Muratorio, S. and Klein, E. K. (2008). Comparing direct vs. indirect estimates of gene flow within a population of a scattered tree species. Molecular Ecology, 17(11), 2743-2754. doi: 10.1111/j.1365-294X.2008.03783.x
Review: 1

Soil variation response is mediated by growth trajectories rather than functional traits in a widespread pioneer Neotropical tree
Growth trajectories, better than organ-level functional traits, reveal intraspecific response to environmental variation
Recommended by François Munoz based on reviews by Georges Kunstler and François MunozFunctional traits are “morpho-physio-phenological traits which impact fitness indirectly via their effects on growth, reproduction and survival” [1]. Most functional traits are defined at organ level, e.g. for leaves, roots and stems, and reflect key aspects of resource acquisition and resource use by organisms for their development and reproduction [2]. More rarely, some functional traits can be related to spatial development, such as vegetative height and lateral spread in plants.
Organ-level traits are especially popular because they can be measured in a standard way and easily compared over many plants. But these traits can broadly vary during the life of an organism. For instance, Roggy et al. [3] found that Leaf Mass Area can vary from 30 to 140 g.m^(-2) between seedling and adult stages for the canopy tree Dicorynia guianensis in French Guiana. Fortunel et al. [4] have also showed that developmental stages much contribute to functional trait variation within several Micropholis tree species in lowland Amazonia.
The way plants grow and invest resources into organs is variable during life and allows defining specific developmental sequences and architectural models [5,6]. There is clear ontogenic variation in leaf number, leaf properties and ramification patterns. Ontogenic variations reflect changing adaptation of an individual over its life, depending on the changing environmental conditions.
In this regard, measuring a single functional trait at organ level in adult trees should miss the variation of resource acquisition and use strategies over time. Thus we should built a more integrative approach of ecological development, also called “eco-devo” approach [7].
Although the ecological significance of ontogeny and developmental strategies is now well known, the extent to which it contributes to explain species survival and coexistence in communities is still broadly ignored in functional ecology. Levionnois et al. [8] investigated intraspecific variation of functional traits and growth trajectories in a typical, early-successional tree species in French Guiana, Amazonia. This species, Cecropia obtusa, is generalist regarding soil type and can be found on both white sand and ferralitic soil. The study examines whether there in intraspecific variation in functional traits and growth trajectories of C. obtusa in response to the contrasted soil types.
The tree communities observed on the two types of soils include species with distinctive functional trait values, that is, there are changes in species composition related to different species strategies along the classical wood and leaf economic spectra. The populations of C. obtusa found on the two soils showed some difference in functional traits, but it did not concern traits related to the main economic spectra. Conversely, the populations showed different growth strategies, in terms of spatial and temporal development.
The major lessons we can learn from the study are:
(i) Functional traits measured at organ level cannot reflect well how long-lived plants collect and invest resources during their life. The results show the potential of considering architectural and developmental traits together with organ-level functional traits, to better acknowledge the variation in ecological strategies over plant life, and thus to better understand community assembly processes.
(ii) What makes functional changes between communities differs when considering interspecific and intraspecific variation. Species turnover should encompass different corteges of soil specialists. These specialists are sorted along economic spectra, as shown in tropical rainforests and globally [2]. Conversely, a generalist species such as C. obtusa does occur on contrasted soil, which entails that it can accommodate the contrasted ecological conditions. However, the phenotypic adjustment is not related to how leaves and wood ensure photosynthesis, water and nutrient acquisition, but regards the way the resources are allocated to growth and reproduction over time.
The results of the study stress the need to better integrate growth strategies and ontogeny in the research agenda of functional ecology. We can anticipate that organ-level functional traits and growth trajectories will be more often considered together in ecological studies. The integration should help better understand the temporal niche of organisms, and how organisms can coexist in space and time with other organisms during their life. Recently, Klimešová et al. [9] have proposed standardized protocols for collecting plant modularity traits. Such effort to propose easy-to-measure traits representing plant development and ontogeny, with clear functional roles, should foster the awaited development of an “eco-devo” approach.
References
[1] Violle, C., Navas, M. L., Vile, D., Kazakou, E., Fortunel, C., Hummel, I., & Garnier, E. (2007). Let the concept of trait be functional!. Oikos, 116(5), 882-892. doi: 10.1111/j.0030-1299.2007.15559.x
[2] Díaz, S. et al. (2016). The global spectrum of plant form and function. Nature, 529(7585), 167-171. doi: 10.1038/nature16489
[3] Roggy, J. C., Nicolini, E., Imbert, P., Caraglio, Y., Bosc, A., & Heuret, P. (2005). Links between tree structure and functional leaf traits in the tropical forest tree Dicorynia guianensis Amshoff (Caesalpiniaceae). Annals of forest science, 62(6), 553-564. doi: 10.1051/forest:2005048
[4] Fortunel, C., Stahl, C., Heuret, P., Nicolini, E. & Baraloto, C. (2020). Disentangling the effects of environment and ontogeny on tree functional dimensions for congeneric species in tropical forests. New Phytologist. doi: 10.1111/nph.16393
[5] Barthélémy, D., & Caraglio, Y. (2007). Plant architecture: a dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. Annals of botany, 99(3), 375-407. doi: 10.1093/aob/mcl260
[6] Hallé, F., & Oldeman, R. A. (1975). An essay on the architecture and dynamics of growth of tropical trees. Kuala Lumpur: Penerbit Universiti Malaya.
[7] Sultan, S. E. (2007). Development in context: the timely emergence of eco-devo. Trends in Ecology & Evolution, 22(11), 575-582. doi: 10.1016/j.tree.2007.06.014
[8] Levionnois, S., Tysklind, N., Nicolini, E., Ferry, B., Troispoux, V., Le Moguedec, G., Morel, H., Stahl, C., Coste, S., Caron, H. & Heuret, P. (2020). Soil variation response is mediated by growth trajectories rather than functional traits in a widespread pioneer Neotropical tree. bioRxiv, 351197, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/351197
[9] Klimešová, J. et al. (2019). Handbook of standardized protocols for collecting plant modularity traits. Perspectives in Plant Ecology, Evolution and Systematics, 40, 125485. doi: 10.1016/j.ppees.2019.125485