Environmental perturbations and transitions between ecological and evolutionary equilibria: an eco-evolutionary feedback framework
Tim Coulson
https://doi.org/10.1101/509067
Stasis and the phenotypic gambit
Recommended by Tom Van Dooren based on reviews by Jacob Johansson, Katja Räsänen and 1 anonymous reviewer
The preprint "Environmental perturbations and transitions between ecological and evolutionary equilibria: an eco-evolutionary feedback framework" by Coulson (2020) presents a general framework for evolutionary ecology, useful to interpret patterns of selection and evolutionary responses to environmental transitions. The paper is written in an accessible and intuitive manner. It reviews important concepts which are at the heart of evolutionary ecology. Together, they serve as a worldview which you can carry with you to interpret patterns in data or observations in nature. I very much appreciate it that Coulson (2020) presents his personal intuition laid bare, the framework he uses for his research and how several strong concepts from theoretical ecology fit in there. Overviews as presented in this paper are important to understand how we as researchers put the pieces together.
A main message of the paper is that resource detection and acquisition traits, broadly called "resource accrual traits" are at the core of evolutionary dynamics. These traits and the processes they are involved in often urge some degree of individual specialization. Not all traits are resource accrual traits all the time. Guppies are cited as an example, which have traits in high predation environments that make foraging easier for them, such as being less conspicuous to predators. In the absence of predators, these same traits might be neutral. Their colour pattern might then contribute much less to the odds of obtaining resources.
"Resource accrual" reminds me of discussions of resource holding potential (Parker 1974), which can be for example the capacity to remain on a bird feeder without being dislodged. However, the idea is much broader and aggression does not need to be important for the acquisition of resources. Evolutionary success is reserved for those steadily obtaining resources. This recalls the pessimization principle of Metz et al. (2008), which applies in a restricted set of situations and where the strategy which persists at the lowest resource levels systematically wins evolutionary contests. If this principle would apply universally, the world then inherently become the worst possible. Resources determine energy budgets and different life history strategies allocate these differently to maximize fitness. The fine grain of environments and the filtration by individual histories generate a lot of variation in outcomes. However, constraint-centered approaches (Kempes et al. 2019, Kooijman 2010) are mentioned but are not at the core of this preprint. Evolution is rather seen as dynamic programming optimization with interactions within and between species. Coulson thus extends life history studies such as for example Tonnabel et al. (2012) with eco-evolutionary feedbacks. Examples used are guppies, algae-rotifer interactions and others. Altogether, this makes for an optimistic paper pushing back the pessimization principle.
Populations are expected to spend most of the time in quasi-equilibrium states where the long run stochastic growth rate is close to zero for all genotypes, alleles or other chosen classes. In the preprint, attention is given to reproductive value calculus, another strong tool in evolutionary dynamics (Grafen 2006, Engen et al. 2009), which tells us how classes within a population contribute to population composition in the distant future. The expected asymptotic fitness of an individual is equated to its expected reproductive value, but this might require particular ways of calculating reproductive values (Coulson 2020). Life history strategies can also be described by per generation measures such as R0 (currently on everyone's radar due to the coronavirus pandemic), generation time etc. Here I might disagree because I believe that this focus in per generation measures can lead to an incomplete characterization of plastic and other strategies involved in strategies such as bet-hedging. A property at quasi-equilibrium states is precise enough to serve as a null hypothesis which can be falsified: all types must in the long run leave equal numbers of descendants. If there is any property in evolutionary ecology which is useful it is this one and it rightfully merits attention.
However, at quasi-equilibrium states, directional selection has been observed, often without the expected evolutionary response. The preprint aims to explain this and puts forward the presence of non-additive gene action as a mechanism. I don't believe that it is the absence of clonal inheritance which matters very much in itself (Van Dooren 2006) unless genes with major effect are present in protected polymorphisms. The preprint remains a bit unclear on how additive gene action is broken, and here I add from the sphere in which I operate. Non-additive gene action can be linked to non-linear genotype-phenotype maps (Van Dooren 2000, Gilchrist and Nijhout 2001) and if these maps are non-linear enough to create constraints on phenotype determination, by means of maximum or minimum phenotypes which cannot be surpassed for any combination of the underlying traits, then they create additional evolutionary quasi-equilibrium states, with directional selection on a phenotype such as body size. I believe Coulson hints at this option (Coulson et al. 2006), but also at a different one: if body size is mostly determined by variation in resource accrual traits, then the resource accrual traits can be under stabilizing selection while body size is not. This requires that all resource accrual traits affect other phenotypic or demographic properties next to body size. In both cases, microevolutionary outcomes cannot be inferred from inspecting body sizes alone, either resource accrual traits need to be included explicitly, or non-linearities, or both when the map between resource accrual and body size is non-linear (Van Dooren 2000).
The discussion of the phenotypic gambit (Grafen 1984) leads to another long-standing issue in evolutionary biology. Can predictions of adaptation be made by inspecting and modelling individual phenotypes alone? I agree that with strongly non-linear genotype-phenotype maps they cannot and for multivariate sets of traits, genetic and phenotypic correlations can be very different (Hadfield et al. 2007). However, has the phenotypic gambit ever claimed to be valid globally or should it rather be used locally for relatively small amounts of variation? Grafen (1984) already contained caveats which are repeated here. As a first approximation, additivity might produce quite correct predictions and thus make the gambit operational in many instances. When important individual traits are omitted, it may just be misspecified. I am interested to see cases where the framework Coulson (2020) proposes is used for very large numbers of phenotypic and genotypic attributes. In the end, these highly dimensional trait distributions might basically collapse to a few major axes of variation due to constraints on resource accrual.
I highly recommend reading this preprint and I am looking forward to the discussion it will generate.
References
[1] Coulson, T. (2020) Environmental perturbations and transitions between ecological and evolutionary equilibria: an eco-evolutionary feedback framework. bioRxiv, 509067, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/509067
[2] Coulson, T., Benton, T. G., Lundberg, P., Dall, S. R. X., and Kendall, B. E. (2006). Putting evolutionary biology back in the ecological theatre: a demographic framework mapping genes to communities. Evolutionary Ecology Research, 8(7), 1155-1171.
[3] Engen, S., Lande, R., Sæther, B. E. and Dobson, F. S. (2009) Reproductive value and the stochastic demography of age-structured populations. The American Naturalist 174: 795-804. doi: 10.1086/647930
[4] Gilchrist, M. A. and Nijhout, H. F. (2001). Nonlinear developmental processes as sources of dominance. Genetics, 159(1), 423-432.
[5] Grafen, A. (1984) Natural selection, kin selection and group selection. In: Behavioural Ecology: An Evolutionary Approach,2nd edn (JR Krebs & NB Davies eds), pp. 62–84. Blackwell Scientific, Oxford.
[6] Grafen, A. (2006). A theory of Fisher's reproductive value. Journal of mathematical biology, 53(1), 15-60. doi: 10.1007/s00285-006-0376-4
[7] Hadfield, J. D., Nutall, A., Osorio, D. and Owens, I. P. F. (2007). Testing the phenotypic gambit: phenotypic, genetic and environmental correlations of colour. Journal of evolutionary biology, 20(2), 549-557. doi: 10.1111/j.1420-9101.2006.01262.x
[8] Kempes, C. P., West, G. B., and Koehl, M. (2019). The scales that limit: the physical boundaries of evolution. Frontiers in Ecology and Evolution, 7, 242. doi: 10.3389/fevo.2019.00242
[9] Kooijman, S. A. L. M. (2010) Dynamic Energy Budget theory for metabolic organisation. University Press, third edition.
[10] Metz, J. A. J., Mylius, S.D. and Diekman, O. (2008) When does evolution optimize?. Evolutionary Ecology Research 10: 629-654.
[11] Parker, G. A. (1974). Assessment strategy and the evolution of fighting behaviour. Journal of theoretical Biology, 47(1), 223-243. doi: 10.1016/0022-5193(74)90111-8
[12] Tonnabel, J., Van Dooren, T. J. M., Midgley, J., Haccou, P., Mignot, A., Ronce, O., and Olivieri, I. (2012). Optimal resource allocation in a serotinous non‐resprouting plant species under different fire regimes. Journal of Ecology, 100(6), 1464-1474. doi: 10.1111/j.1365-2745.2012.02023.x
[13] Van Dooren, T. J. M. (2000). The evolutionary dynamics of direct phenotypic overdominance: emergence possible, loss probable. Evolution, 54(6), 1899-1914. doi: 10.1111/j.0014-3820.2000.tb01236.x
[14] Van Dooren, T. J. M. (2006). Protected polymorphism and evolutionary stability in pleiotropic models with trait‐specific dominance. Evolution, 60(10), 1991-2003. doi: 10.1111/j.0014-3820.2006.tb01837.x
| Environmental perturbations and transitions between ecological and evolutionary equilibria: an eco-evolutionary feedback framework | Tim Coulson | <p>I provide a general framework for linking ecology and evolution. I start from the fact that individuals require energy, trace molecules, water, and mates to survive and reproduce, and that phenotypic resource accrual traits determine an individ... | 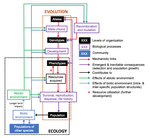 | Eco-evolutionary dynamics, Evolutionary ecology | Tom Van Dooren | | 2019-01-03 10:05:16 | View |
Which pitfall traps and sampling efforts should be used to evaluate the effects of cropping systems on the taxonomic and functional composition of arthropod communities?
Antoine Gardarin and Muriel Valantin-Morison
https://doi.org/10.5281/zenodo.3468920
On the importance of experimental design: pitfall traps and arthropod communities
Recommended by Ignasi Bartomeus based on reviews by Cécile ALBERT and Matthias Foellmer based on reviews by Cécile ALBERT and Matthias Foellmer
Despite the increasing refinement of statistical methods, a robust experimental design is still one of the most important cornerstones to answer ecological and evolutionary questions. However, there is a strong trade-off between a perfect design and its feasibility. A common mantra is that more data is always better, but how much is enough is complex to answer, specially when we want to capture the spatial and temporal variability of a given process. Gardarin and Valantin-Morison [1] make an effort to answer these questions for a practical case: How many pitfalls traps, of which type, and over which extent, do we need to detect shifts in arthropod community composition in agricultural landscapes. There is extense literature on how to approach these challenges using preliminary data in combination with simulation methods [e.g. 2], but practical cases are always welcomed to illustrate the complexity of the decisions to be made. A key challenge in this situation is the nature of simplified and patchy agricultural arthropod communities. In this context, small effect sizes are expected, but those small effects are relevant from an ecological point of view because small increases at low biodiversity may produce large gains in ecosystem functioning [3].
The paper shows that some variables are not important, such as the type of fluid used to fill the pitfall traps. This is good news for potential comparisons among studies using slightly different protocols. However, the bad news are that the sampling effort needed for detecting community changes is larger than the average effort currently implemented. A potential solution is to focus on Community Weighed Mean metrics (CWM; i.e. a functional descriptor of the community body size distribution) rather than on classic metrics such as species richness, as detecting changes on CWM requires a lower sampling effort and it has a clear ecological interpretation linked to ecosystem functioning.
Beyond the scope of the data presented, which is limited to a single region over two years, and hence it is hard to extrapolate to other regions and years, the big message of the paper is the need to incorporate statistical power simulations as a central piece of the ecologist's toolbox. This is challenging, especially when you face questions such as: Should I replicate over space, or over time? The recommended paper is accompanied by the statistical code used, which should facilitate this task to other researchers. Furthermore, we should be aware that some important questions in ecology are highly variable in space and time, and hence, larger sampling effort across space and time is needed to detect patterns. Larger and longer monitoring schemes require a large effort (and funding), but if we want to make relevant ecology, nobody said it would be easy.
References
[1] Gardarin, A. and Valantin-Morison, M. (2019). Which pitfall traps and sampling efforts should be used to evaluate the effects of cropping systems on the taxonomic and functional composition of arthropod communities? Zenodo, 3468920, ver. 3 peer-reviewed and recommended by PCI Ecology. doi: 10.5281/zenodo.3468920
[2] Johnson, P. C., Barry, S. J., Ferguson, H. M., and Müller, P. (2015). Power analysis for generalized linear mixed models in ecology and evolution. Methods in ecology and evolution, 6(2), 133-142. doi: 10.1111/2041-210X.12306
[3] Cardinale, B. J. et al. (2012). Biodiversity loss and its impact on humanity. Nature, 486(7401), 59-67. doi: 10.1038/nature11148
| Which pitfall traps and sampling efforts should be used to evaluate the effects of cropping systems on the taxonomic and functional composition of arthropod communities? | Antoine Gardarin and Muriel Valantin-Morison | <p>1. Ground dwelling arthropods are affected by agricultural practices, and analyses of their responses to different crop management are required. The sampling efficiency of pitfall traps has been widely studied in natural ecosystems. In arable a... |  | Agroecology, Biodiversity, Biological control, Community ecology | Ignasi Bartomeus | | 2019-01-08 09:40:14 | View |
Community size affects the signals of ecological drift and selection on biodiversity
Tadeu Siqueira, Victor S. Saito, Luis M. Bini, Adriano S. Melo, Danielle K. Petsch, Victor L. Landeiro, Kimmo T. Tolonen, Jenny Jyrkänkallio-Mikkola, Janne Soininen, Jani Heino
https://doi.org/10.1101/515098
Toward an empirical synthesis on the niche versus stochastic debate
Recommended by Eric Harvey based on reviews by Kevin Cazelles and Romain Bertrand
As far back as Clements [1] and Gleason [2], the historical schism between deterministic and stochastic perspectives has divided ecologists. Deterministic theories tend to emphasize niche-based processes such as environmental filtering and species interactions as the main drivers of species distribution in nature, while stochastic theories mainly focus on chance colonization, random extinctions and ecological drift [3]. Although the old days when ecologists were fighting fiercely over null models and their adequacy to capture niche-based processes is over [4], the ghost of that debate between deterministic and stochastic perspectives came back to haunt ecologists in the form of the ‘environment versus space’ debate with the development of metacommunity theory [5]. While interest in that question led to meaningful syntheses of metacommunity dynamics in natural systems [6], it also illustrated how context-dependant the answer was [7]. One of the next frontiers in metacommunity ecology is to identify the underlying drivers of this observed context-dependency in the relative importance of ecological processus [7, 8].
Reflecting on seminal work by Robert MacArthur emphasizing different processes at different spatial scales [9, 10] (the so-called ‘MacArthur paradox’), Chase and Myers proposed in 2011 that a key in solving the deterministic versus stochastic debate was probably to turn our attention to how the relative importance of local processes changes across spatial scales [3]. Scale-dependance is a well-acknowledged challenge in ecology, hampering empirical syntheses and comparisons between studies [11-14]. Embracing the scale-dependance of ecological processes would not only lead to stronger syntheses and consolidation of current knowledge, it could also help resolve many current debates or apparent contradictions [11, 15, 16].
The timely study by Siqueira et al. [17] fits well within this historical context by exploring the relative importance of ecological drift and selection across a gradient of community size (number of individuals in a given community). More specifically, they tested the hypothesis that small communities are more dissimilar among each other because of ecological drift compared to large communities, which are mainly structured by niche selection [17]. That smaller populations or communities should be more affected by drift is a mathematical given [18], but the main questions are i) for a given community size how important is ecological drift relative to other processes, and ii) how small does a community have to be before random assembly dominates? The authors answer these questions using an extensive stream dataset with a community size gradient sampled from 200 streams in two climatic regions (Brazil and Finland). Combining linear models with recent null model approaches to measure deviations from random expectations [19], they show that, as expected based on theory and recent experimental work, smaller communities tend to have higher β-diversity, and that those β-diversity patterns could not be distinguished from random assembly processes [17]. Spatial turnover among larger communities is mainly driven by niche-based processes related to species sorting or dispersal dynamics [17]. Given the current environmental context, with many anthropogenic perturbations leading to reduced community size, it is legitimate to wonder, as the authors do, whether we are moving toward a more stochastic and thus less predictable world with obvious implications for the conservation of biodiversity [17].
The real strength of the study by Siqueira et al. [17], in my opinion, is in the inclusion of stream data from boreal and tropical regions. Interestingly and most importantly, the largest communities in the tropical streams are as large as the smallest communities in the boreal streams. This is where the study should really have us reflect on the notions of context-dependency in observed patterns because the negative relationship between community size and β-diversity was only observed in the tropical streams, but not in the boreal streams [17]. This interesting nonlinearity in the response means that a study that would have investigated the drift versus niche-based question only in Finland would have found very different results from the same study in Brazil. Only by integrating such a large scale gradient of community sizes together could the authors show the actual shape of the relationship, which is the first step toward building a comprehensive synthesis on a debate that has challenged ecologists for almost a century.
References
[1] Clements, F. E. (1936). Nature and structure of the climax. Journal of ecology, 24(1), 252-284. doi: 10.2307/2256278
[2] Gleason, H. A. (1917). The structure and development of the plant association. Bulletin of the Torrey Botanical Club, 44(10), 463-481. doi: 10.2307/2479596
[3] Chase, J. M., and Myers, J. A. (2011). Disentangling the importance of ecological niches from stochastic processes across scales. Philosophical transactions of the Royal Society B: Biological sciences, 366(1576), 2351-2363. doi: 10.1098/rstb.2011.0063
[4] Diamond, J. M., and Gilpin, M. E. (1982). Examination of the “null” model of Connor and Simberloff for species co-occurrences on islands. Oecologia, 52(1), 64-74. doi: 10.1007/BF00349013
[5] Leibold M. A., et al. (2004). The metacommunity concept: a framework for multi‐scale community ecology. Ecology letters, 7(7), 601-613. doi: 10.1111/j.1461-0248.2004.00608.x
[6] Cottenie, K. (2005). Integrating environmental and spatial processes in ecological community dynamics. Ecology letters, 8(11), 1175-1182. doi: 10.1111/j.1461-0248.2005.00820.x
[7] Leibold, M. A. and Chase, J. M. (2018). Metacommunity Ecology. Monographs in Population Biology, vol. 59. Princeton University Press.
[8] Vellend, M. (2010). Conceptual synthesis in community ecology. The Quarterly review of biology, 85(2), 183-206. doi: 10.1086/652373
[9] MacArthur, R. H., and Wilson, E. O. (1963). An equilibrium theory of insular zoogeography. Evolution, 17(4), 373-387. doi: 10.1111/j.1558-5646.1963.tb03295.x
[10] MacArthur, R. H., and Levins, R. (1967). The limiting similarity, convergence, and divergence of coexisting species. The American Naturalist, 101(921), 377-385. doi: 10.1086/282505
[11] Viana, D. S., and Chase, J. M. (2019). Spatial scale modulates the inference of metacommunity assembly processes. Ecology, 100(2), e02576. doi: 10.1002/ecy.2576
[12] Chave, J. (2013). The problem of pattern and scale in ecology: what have we learned in 20 years?. Ecology letters, 16, 4-16. doi: 10.1111/ele.12048
[13] Patrick, C. J., and Yuan, L. L. (2019). The challenges that spatial context present for synthesizing community ecology across scales. Oikos, 128(3), 297-308. doi: 10.1111/oik.05802
[14] Chase, J. M., and Knight, T. M. (2013). Scale‐dependent effect sizes of ecological drivers on biodiversity: why standardised sampling is not enough. Ecology letters, 16, 17-26. doi: 10.1111/ele.12112
[15] Horváth, Z., Ptacnik, R., Vad, C. F., and Chase, J. M. (2019). Habitat loss over six decades accelerates regional and local biodiversity loss via changing landscape connectance. Ecology letters, 22(6), 1019-1027. doi: 10.1111/ele.13260
[16] Chase, J. M, Gooriah, L., May, F., Ryberg, W. A, Schuler, M. S, Craven, D., and Knight, T. M. (2019). A framework for disentangling ecological mechanisms underlying the island species–area relationship. Frontiers of Biogeography, 11(1). doi: 10.21425/F5FBG40844.
[17] Siqueira T., Saito V. S., Bini L. M., Melo A. S., Petsch D. K. , Landeiro V. L., Tolonen K. T., Jyrkänkallio-Mikkola J., Soininen J. and Heino J. (2019). Community size affects the signals of ecological drift and niche selection on biodiversity. bioRxiv 515098, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/515098
[18] Hastings A., Gross L. J. eds. (2012). Encyclopedia of theoretical ecology (University of California Press, Berkeley).
[19] Chase, J. M., Kraft, N. J., Smith, K. G., Vellend, M., and Inouye, B. D. (2011). Using null models to disentangle variation in community dissimilarity from variation in α‐diversity. Ecosphere, 2(2), 1-11. doi: 10.1890/ES10-00117.1
| Community size affects the signals of ecological drift and selection on biodiversity | Tadeu Siqueira, Victor S. Saito, Luis M. Bini, Adriano S. Melo, Danielle K. Petsch, Victor L. Landeiro, Kimmo T. Tolonen, Jenny Jyrkänkallio-Mikkola, Janne Soininen, Jani Heino | <p>Ecological drift can override the effects of deterministic niche selection on small populations and drive the assembly of small communities. We tested the hypothesis that smaller local communities are more dissimilar among each other because of... |  | Biodiversity, Coexistence, Community ecology, Competition, Conservation biology, Dispersal & Migration, Freshwater ecology, Spatial ecology, Metacommunities & Metapopulations | Eric Harvey | | 2019-01-09 19:06:21 | View |
Does phenology explain plant-pollinator interactions at different latitudes? An assessment of its explanatory power in plant-hoverfly networks in French calcareous grasslands
Natasha de Manincor, Nina Hautekeete, Yves Piquot, Bertrand Schatz, Cédric Vanappelghem, François Massol
https://doi.org/10.5281/zenodo.2543768
The role of phenology for determining plant-pollinator interactions along a latitudinal gradient
Recommended by Anna Eklöf based on reviews by Ignasi Bartomeus, Phillip P.A. Staniczenko and 1 anonymous reviewer
Increased knowledge of what factors are determining species interactions are of major importance for our understanding of dynamics and functionality of ecological communities [1]. Currently, when ongoing temperature modifications lead to changes in species temporal and spatial limits the subject gets increasingly topical. A species phenology determines whether it thrive or survive in its environment. However, as the phenologies of different species are not necessarily equally affected by environmental changes, temporal or spatial mismatches can occur and affect the species-species interactions in the network [2] and as such the full network structure.
In this preprint by Manincor et al. [3] the authors explore the effect of phenology overlap on a large network of species interactions in calcareous grasslands in France. They analyze if and how this effect varies along a latitudinal gradient using empirical data on six plant-hoverfly networks. When comparing ecological network along gradients a well-known problem is that the network metrics is dependent on network size [4]. Therefore, instead of focusing on complete network structure the authors here focus on the factors that determine the probability of interactions and interaction frequency (number of visits). The authors use Bayesian Structural Equation Models (SEM) to link the interaction probability and number of visits to phenology overlap and species abundance. SEM is a multivariate technique that can be used to test several hypotheses and evaluate multiple causal relationships using both observed and latent variables to explain some other observed variables. The authors provide a nice description of the approach for this type of study system. In addition, the study also tests whether phenology affects network compartmentalization, by analyzing species subgroups using a latent block model (LBM) which is a clustering method particularly well-suited for weighted networks.
The authors identify phenology overlap as an important determinant of plant-pollinator interactions, but also conclude this factor alone is not sufficient to explain the species interactions. Species abundances was important for number of visits. Plant phenology drives the duration of the phenology overlap between plant and hoverflies in the studied system. This in turn influences either the probability of interaction or the expected number of visits, as well as network compartmentalization. Longer phenologies correspond to lower modularity inferring less constrained interactions, and shorter phenologies correspond to higher modularity inferring more constrained interactions.
What make this study particularly interesting is the presentation of SEMs as an innovative approach to compare networks of different sizes along environmental gradients. The authors show that these methods can be a useful tool when the aim is to understand the structure of plant-pollinator networks and data is varying in complexities. During the review process the authors carefully addressed to the comments from the two reviewers and the manuscript improved during the process. Both reviewers have expertise highly relevant for the research performed and the development of the manuscript. In my opinion this is a highly interesting and valuable piece of work both when it comes to the scientific question and the methodology. I look forward to further follow this research.
References
[1] Pascual, M., and Dunne, J. A. (Eds.). (2006). Ecological networks: linking structure to dynamics in food webs. Oxford University Press.
[2] Parmesan, C. (2007). Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Global Change Biology, 13(9), 1860-1872. doi: 10.1111/j.1365-2486.2007.01404.x
[3] de Manincor, N., Hautekeete, N., Piquot, Y., Schatz, B., Vanappelghem, C. and Massol, F. (2019). Does phenology explain plant-pollinator interactions at different latitudes? An assessment of its explanatory power in plant-hoverfly networks in French calcareous grasslands. Zenodo, 2543768, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.5281/zenodo.2543768
[4] Staniczenko, P. P., Kopp, J. C., and Allesina, S. (2013). The ghost of nestedness in ecological networks. Nature communications, 4, 1391. doi: 10.1038/ncomms2422
| Does phenology explain plant-pollinator interactions at different latitudes? An assessment of its explanatory power in plant-hoverfly networks in French calcareous grasslands | Natasha de Manincor, Nina Hautekeete, Yves Piquot, Bertrand Schatz, Cédric Vanappelghem, François Massol | <p>For plant-pollinator interactions to occur, the flowering of plants and the flying period of pollinators (i.e. their phenologies) have to overlap. Yet, few models make use of this principle to predict interactions and fewer still are able to co... |  | Interaction networks, Pollination, Statistical ecology | Anna Eklöf | | 2019-01-18 19:02:13 | View |
A community perspective on the concept of marine holobionts: current status, challenges, and future directions
Simon M. Dittami, Enrique Arboleda, Jean-Christophe Auguet, Arite Bigalke, Enora Briand, Paco Cárdenas, Ulisse Cardini, Johan Decelle, Aschwin Engelen, Damien Eveillard, Claire M.M. Gachon, Sarah Griffiths, Tilmann Harder, Ehsan Kayal, Elena Kazamia, Francois H. Lallier, Mónica Medina, Ezequiel M. Marzinelli, Teresa Morganti, Laura Núñez Pons, Soizic Pardo, José Pintado Valverde, Mahasweta Saha, Marc-André Selosse, Derek Skillings, Willem Stock, Shinichi Sunagawa, Eve Toulza, Alexey Vorobev, Cat...
10.5281/zenodo.3696771
Marine holobiont in the high throughput sequencing era
Recommended by Sophie Arnaud-Haond and Corinne Vacher based on reviews by Sophie Arnaud-Haond and Aurélie Tasiemski
The concept of holobiont dates back to more than thirty years, it was primarily coined to hypothesize the importance of symbiotic associations to generate significant evolutionary novelties. Quickly adopted to describe the now well-studied system formed by zooxanthella associated corals, this concept expanded much further after the emergence of High-Throughput Sequencing and associated progresses in metabarcoding and metagenomics.
Holobionts – defined as the association between an individual host and its microbiota - are now increasingly described at sea and on land. The opinion article by Dittami et al. [1] provides a synthetic overview of marine holobionts. It retraces the history of the holobiont concept, recalls the main mechanisms underlying the association between hosts and microbial communities, highlights the influence of these symbioses on marine ecosystem functioning, and outlines current tools and future lines of research.
In particular, the article discusses some particularities of marine systems, such as the strong connectivity allowing an exchange of microorganisms and chemical signals between and within holobionts.
The authors advocate the need to bridge the gap between large scale exploration studies and smaller scale mechanistic studies, by conducting interdisciplinary research (combining physiology, biochemistry, ecology, experimentation and computational modeling) on some keystone holobionts.
Finally, one strength of the paper by Dittami et al. [1] is that it places the concept of the holobiont in an applied research framework. Several possible applications of knowledge on host-microbiota interactions are suggested, both in the field of aquaculture and that of monitoring the health of marine ecosystems. This article contains all the necessary elements for someone who would like to jump into the study of the holobionths in the marine world.
References
[1] Dittami SM, Arboleda E, Auguet J, Bigalke A, Briand E, Cardenas P, Cardini U, Decelle J, Engelen AH, Eveillard D, Gachon CMM, Griffiths SM, Harder T, Kayal E, Kazamia E, Lallier FH, Medina M, Marzinelli E, Morganti T, Núñez Pons L, Prado S, Pintado J, Saha M, Selosse M, Skillings D, Stock W, Sunagawa S, Toulza E, Vorobev A, Leblanc C, Not F. (2020). A community perspective on the concept of marine holobionts: current status, challenges, and future directions. Zenodo, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.5281/zenodo.3696771
| A community perspective on the concept of marine holobionts: current status, challenges, and future directions | Simon M. Dittami, Enrique Arboleda, Jean-Christophe Auguet, Arite Bigalke, Enora Briand, Paco Cárdenas, Ulisse Cardini, Johan Decelle, Aschwin Engelen, Damien Eveillard, Claire M.M. Gachon, Sarah Griffiths, Tilmann Harder, Ehsan Kayal, Elena Kazam... | Host-microbe interactions play crucial roles in marine ecosystems. However, we still have very little understanding of the mechanisms that govern these relationships, the evolutionary processes that shape them, and their ecological consequences. T... | 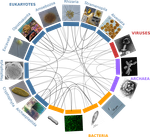 | Marine ecology, Microbial ecology & microbiology, Symbiosis | Sophie Arnaud-Haond | | 2019-02-05 17:57:11 | View |
Interplay between historical and current features of the cityscape in shaping the genetic structure of the house mouse (Mus musculus domesticus) in Dakar (Senegal, West Africa)
Claire Stragier, Sylvain Piry, Anne Loiseau, Mamadou Kane, Aliou Sow, Youssoupha Niang, Mamoudou Diallo, Arame Ndiaye, Philippe Gauthier, Marion Borderon, Laurent Granjon, Carine Brouat, Karine Berthier
https://doi.org/10.1101/557066
Urban past predicts contemporary genetic structure in city rats
Recommended by Michelle DiLeo based on reviews by Torsti Schulz, ? and 1 anonymous reviewer
Urban areas are expanding worldwide, and have become a dominant part of the landscape for many species. Urbanization can fragment pre-existing populations of vulnerable species leading to population declines and the loss of connectivity. On the other hand, expansion of urban areas can also facilitate the spread of human commensals including pests. Knowledge of the features of cityscapes that facilitate gene flow and maintain diversity of pests is thus key to their management and eradication.
Cities are complex mosaics of natural and manmade surfaces, and habitat quality is not only influenced by physical aspects of the cityscape but also by socioeconomic factors and human behaviour. Constant development means that cities also change rapidly in time; contemporary urban life reflects only a snapshot of the environmental conditions faced by populations. It thus remains a challenge to identify the features that actually drive ecology and evolution of populations in cities [1]. While several studies have highlighted strong urban clines in genetic structure and adaption [2], few have considered the influence of factors beyond physical aspects of the cityscape or historical processes.
In this paper, Stragier et al. [3] sought to identify the current and past features of the cityscape and socioeconomic factors that shape genetic structure and diversity of the house mouse (Mus musculus domesticus) in Dakar, Senegal. The authors painstakingly digitized historical maps of Dakar from the time of European settlement in 1862 to present. The authors found that the main spatial genetic cline was best explained by historical cityscape features, with higher apparent gene flow and genetic diversity in areas that were connected earlier to initial European settlements. Beyond the main trend of spatial genetic structure, they found further evidence that current features of the cityscape were important. Specifically, areas with low vegetation and poor housing conditions were found to support large, genetically diverse populations. The authors demonstrate that their results are reproducible using several statistical approaches, including modeling that explicitly accounts for spatial autocorrelation.
The work of Stragier et al. [3] thus highlights that populations of city-dwelling species are the product of both past and present cityscapes. Going forward, urban evolutionary ecologists should consider that despite the potential for rapid evolution in urban landscapes, the signal of a species’ colonization can remain for generations.
References
[1] Rivkin, L. R., Santangelo, J. S., Alberti, M. et al. (2019). A roadmap for urban evolutionary ecology. Evolutionary Applications, 12(3), 384-398. doi: 10.1111/eva.12734
[2] Miles, L. S., Rivkin, L. R., Johnson, M. T., Munshi‐South, J. and Verrelli, B. C. (2019). Gene flow and genetic drift in urban environments. Molecular ecology, 28(18), 4138-4151. doi: 10.1111/mec.15221
[3] Stragier, C., Piry, S., Loiseau, A., Kane, M., Sow, A., Niang, Y., Diallo, M., Ndiaye, A., Gauthier, P., Borderon, M., Granjon, L., Brouat, C. and Berthier, K. (2020). Interplay between historical and current features of the cityscape in shaping the genetic structure of the house mouse (Mus musculus domesticus) in Dakar (Senegal, West Africa). bioRxiv, 557066, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/557066
| Interplay between historical and current features of the cityscape in shaping the genetic structure of the house mouse (Mus musculus domesticus) in Dakar (Senegal, West Africa) | Claire Stragier, Sylvain Piry, Anne Loiseau, Mamadou Kane, Aliou Sow, Youssoupha Niang, Mamoudou Diallo, Arame Ndiaye, Philippe Gauthier, Marion Borderon, Laurent Granjon, Carine Brouat, Karine Berthier | <p>Population genetic approaches may be used to investigate dispersal patterns of species living in highly urbanized environment in order to improve management strategies for biodiversity conservation or pest control. However, in such environment,... | 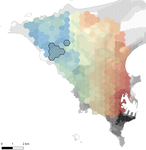 | Biological invasions, Landscape ecology, Molecular ecology | Michelle DiLeo | | 2019-02-22 08:36:13 | View |
Three points of consideration before testing the effect of patch connectivity on local species richness: patch delineation, scaling and variability of metrics
F. Laroche, M. Balbi, T. Grébert, F. Jabot & F. Archaux
https://doi.org/10.1101/640995
Good practice guidelines for testing species-isolation relationships in patch-matrix systems
Recommended by Damaris Zurell based on reviews by 3 anonymous reviewers
Conservation biology is strongly rooted in the theory of island biogeography (TIB). In island systems where the ocean constitutes the inhospitable matrix, TIB predicts that species richness increases with island size as extinction rates decrease with island area (the species-area relationship, SAR), and species richness increases with connectivity as colonisation rates decrease with island isolation (the species-isolation relationship, SIR)[1]. In conservation biology, patches of habitat (habitat islands) are often regarded as analogous to islands within an unsuitable matrix [2], and SAR and SIR concepts have received much attention as habitat loss and habitat fragmentation are increasingly threatening biodiversity [3,4].
The existence of SAR in patch-matrix systems has been confirmed in several studies, while the relative importance of SIR remains debated [2,5] and empirical evidence is mixed. For example, Thiele et al. [6] showed that connectivity effects are trait specific and more important to explain species richness of short-distant dispersers and of specialist species for which the matrix is less permeable. Some authors have also cautioned that the relative support for or against the existence of SIR may depend on methodological decisions related to connectivity metrics, patch classification, scaling decisions and sample size [7].
In this preprint, Laroche and colleagues [8] argue that methodological limits should be fully understood before questioning the validity of SIR in patch-matrix systems. In consequence, they used a virtual ecologist approach [9] to qualify different methodological aspects and derive good practice guidelines related to patch delineation, patch connectivity indices, and scaling of indices with species dispersal distance.
Laroche et al. [8] simulated spatially-explicit neutral meta-communities with up to 100 species in artificial fractal (patch-matrix) landscapes. Each habitat cell could hold up to 100 individuals. In each time step, some individuals died and were replaced by an individual from the regional species pool depending on relative local and regional abundance as well as dispersal distance to the nearest source habitat cell. Different scenarios were run with varying degrees of spatial autocorrelation in the fractal landscape (determining the clumpiness of habitat cells), the proportion of suitable habitat, and the species dispersal distances (with all species showing the same dispersal distance). Laroche and colleagues then sampled species richness in the simulated meta-communities, computed different local connectivity indices for the simulated landscapes (Buffer index with different radii, dIICflux index and dF index, and, finally, related species richness to connectivity.
The complex simulations allowed Laroche and colleagues [8] to test how methodological choices and landscape features may affect SIR. Overall, they found that patch delineation is crucial and should be fine enough to exclude potential within-patch dispersal limitations, and the scaling of the connectivity indices (in simplified words, the window of analyses) should be tailored to the dispersal distance of the species group. Of course, tuning the scaling parameters will be more complicated when dispersal distances vary across species but overall these results corroborate empirical findings that SIR effects are trait specific [6]. Additionally, the results by Laroche and colleagues [8] indicated that indices based on Euclidian rather than topological distance are more performant and that evidence of SIR is more likely if Buffer indices are highly variable between sampled patches.
Although the study is very technical due to the complex simulation approach and the different methods tested, I hope it will not only help guiding methodological choices but also inspire ecologists to further test or even revisit SIR (and SAR) hypotheses for different systems. Also, Laroche and colleagues propose many interesting avenues that could still be explored in this context, for example determining the optimal grid resolution for the patch delineation in empirical studies.
References
[1] MacArthur, R.H. and Wilson, E.O. (1967) The theory of island biogeography. Princeton University Press, Princeton.
[2] Fahrig, L. (2013) Rethinking patch size and isolation effects: the habitat amount hypothesis. Journal of Biogeography, 40(9), 1649-1663. doi: 10.1111/jbi.12130
[3] Hanski, I., Zurita, G.A., Bellocq, M.I. and Rybicki J (2013) Species–fragmented area relationship. Proceedings of the National Academy of Sciences U.S.A., 110(31), 12715-12720. doi: 10.1073/pnas.1311491110
[4] Giladi, I., May, F., Ristow, M., Jeltsch, F. and Ziv, Y. (2014) Scale‐dependent species–area and species–isolation relationships: a review and a test study from a fragmented semi‐arid agro‐ecosystem. Journal of Biogeography, 41(6), 1055-1069. doi: 10.1111/jbi.12299
[5] Hodgson, J.A., Moilanen, A., Wintle, B.A. and Thomas, C.D. (2011) Habitat area, quality and connectivity: striking the balance for efficient conservation. Journal of Applied Ecology, 48(1), 148-152. doi: 10.1111/j.1365-2664.2010.01919.x
[6] Thiele, J., Kellner, S., Buchholz, S., and Schirmel, J. (2018) Connectivity or area: what drives plant species richness in habitat corridors? Landscape Ecology, 33, 173-181. doi: 10.1007/s10980-017-0606-8
[7] Vieira, M.V., Almeida-Gomes, M., Delciellos, A.C., Cerqueira, R. and Crouzeilles, R. (2018) Fair tests of the habitat amount hypothesis require appropriate metrics of patch isolation: An example with small mammals in the Brazilian Atlantic Forest. Biological Conservation, 226, 264-270. doi: 10.1016/j.biocon.2018.08.008
[8] Laroche, F., Balbi, M., Grébert, T., Jabot, F. and Archaux, F. (2020) Three points of consideration before testing the effect of patch connectivity on local species richness: patch delineation, scaling and variability of metrics. bioRxiv, 640995, ver. 5 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/640995
[9] Zurell, D., Berger, U., Cabral, J.S., Jeltsch, F., Meynard, C.N., Münkemüller, T., Nehrbass, N., Pagel, J., Reineking, B., Schröder, B. and Grimm, V. (2010) The virtual ecologist approach: simulating data and observers. Oikos, 119(4), 622-635. doi: 10.1111/j.1600-0706.2009.18284.x
| Three points of consideration before testing the effect of patch connectivity on local species richness: patch delineation, scaling and variability of metrics | F. Laroche, M. Balbi, T. Grébert, F. Jabot & F. Archaux | <p>The Theory of Island Biogeography (TIB) promoted the idea that species richness within sites depends on site connectivity, i.e. its connection with surrounding potential sources of immigrants. TIB has been extended to a wide array of fragmented... | 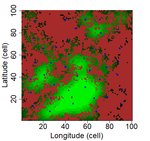 | Biodiversity, Community ecology, Dispersal & Migration, Landscape ecology, Spatial ecology, Metacommunities & Metapopulations | Damaris Zurell | | 2019-05-20 16:03:47 | View |
Comparing statistical and mechanistic models to identify the drivers of mortality within a rear-edge beech population
Cathleen Petit-Cailleux, Hendrik Davi, François Lefevre, Christophe Hurson, Joseph Garrigue, Jean-André Magdalou, Elodie Magnanou and Sylvie Oddou-Muratorio
https://doi.org/10.1101/645747
The complexity of predicting mortality in trees
Recommended by Lucía DeSoto based on reviews by Lisa Hülsmann and 2 anonymous reviewers
One of the main issues of forest ecosystems is rising tree mortality as a result of extreme weather events (Franklin et al., 1987). Eventually, tree mortality reduces forest biomass (Allen et al., 2010), although its effect on forest ecosystem fluxes seems not lasting too long (Anderegg et al., 2016). This controversy about the negative consequences of tree mortality is joined to the debate about the drivers triggering and the mechanisms accelerating tree decline. For instance, there is still room for discussion about carbon starvation or hydraulic failure determining the decay processes (Sevanto et al., 2014) or about the importance of mortality sources (Reichstein et al., 2013). Therefore, understanding and predicting tree mortality has become one of the challenges for forest ecologists in the last decade, doubling the rate of articles published on the topic (*). Although predicting the responses of ecosystems to environmental change based on the traits of species may seem a simplistic conception of ecosystem functioning (Sutherland et al., 2013), identifying those traits that are involved in the proneness of a tree to die would help to predict how forests will respond to climate threatens.
Modelling tree mortality is complex, involving multiple factors acting simultaneously at different scales, from tree genetics to ecosystem dynamics and from microsite conditions to global climatic events. Therefore, taking into account different approaches to reduce uncertainty of the predictions is needed (Bugmann et al., 2019). Petit-Cailleux et al. (2020) uses statistical and process-based models to detect the main mortality drivers of a drought- and frost-prone beech population. Particularly, they assessed the intra-individual characteristics of the population, that may play a decisive role explaining the differences in tree vulnerability to extreme weather events. Comparing the results of both analytical approaches, they find out several key factors, such as defoliation, leaf phenology and tree size, that were consistent between them. Even more, the process-based model showed the physiological mechanisms that may explain the individual vulnerability, for instance higher loss of hydraulic conductance may increase the mortality risk of trees with early budburst phenology and large stem diameter. The authors also successfully model annual mortality rate with a linear relationship including only three parameters: loss of conductance, biomass of reserves and late frost days.
This valuable study is a good example of the complexity in understanding and predicting tree mortality. The authors carried out the ambitious commitment of studying the inter-annual variation in mortality with 14-year dataset. However, it might be not enough time to control for the dependence of temporal data to soundly model mortality rate. The authors also acknowledge that the use of two approaches increases the knowledge from different perspectives, but at the same time comparing their results is difficult because the parameters used are not identical. Particularly, process-based models tend to consider the same microclimatic conditions for every tree in the population, and may produce inconsistences with statistical models. Alternatively, individual-based modelling might overcome some of the incompatibilities between the approaches (Zhu et al., 2019).
(*) Number (and percentage) of articles found in Web of Sciences after searching (December the 10th, 2020) “tree mortality”: from 163 (0.006%) in 2010 to 412 (0.013%) in 2020.
References
Allen et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest ecology and management, 259(4), 660-684. doi: https://doi.org/10.1016/j.foreco.2009.09.001
Anderegg et al. (2016). When a tree dies in the forest: scaling climate-driven tree mortality to ecosystem water and carbon fluxes. Ecosystems, 19(6), 1133-1147. doi: https://doi.org/10.1007/s10021-016-9982-1
Bugmann et al. (2019). Tree mortality submodels drive simulated long‐term forest dynamics: assessing 15 models from the stand to global scale. Ecosphere, 10(2), e02616. doi: https://doi.org/10.1002/ecs2.2616
Franklin, J. F., Shugart, H. H. and Harmon, M. E. (1987) Death as an ecological process: the causes, consequences, and variability of tree mortality. BioScience, 37, 550–556. doi: https://doi.org/10.2307/1310665
Petit-Cailleux, C., Davi, H., Lefèvre, F., Garrigue, J., Magdalou, J.-A., Hurson, C., Magnanou, E. and Oddou-Muratorio, S. (2020) Comparing statistical and mechanistic models to identify the drivers of mortality within a rear-edge beech population. bioRxiv, 645747, ver 7 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/645747
Reichstein et al. (2013). Climate extremes and the carbon cycle. Nature, 500(7462), 287-295. doi: https://doi.org/10.1038/nature12350
Sevanto, S., Mcdowell, N. G., Dickman, L. T., Pangle, R., and Pockman, W. T. (2014). How do trees die? A test of the hydraulic failure and carbon starvation hypotheses. Plant, cell & environment, 37(1), 153-161. doi: https://doi.org/10.1111/pce.12141
Sutherland et al. (2013). Identification of 100 fundamental ecological questions. Journal of ecology, 101(1), 58-67. doi: https://doi.org/10.1111/1365-2745.12025
Zhu, Y., Liu, Z., and Jin, G. (2019). Evaluating individual-based tree mortality modeling with temporal observation data collected from a large forest plot. Forest Ecology and Management, 450, 117496. doi: https://doi.org/10.1016/j.foreco.2019.117496
| Comparing statistical and mechanistic models to identify the drivers of mortality within a rear-edge beech population | Cathleen Petit-Cailleux, Hendrik Davi, François Lefevre, Christophe Hurson, Joseph Garrigue, Jean-André Magdalou, Elodie Magnanou and Sylvie Oddou-Muratorio | <p>Since several studies have been reporting an increase in the decline of forests, a major issue in ecology is to better understand and predict tree mortality. The interactions between the different factors and the physiological processes giving ... |  | Climate change, Physiology, Population ecology | Lucía DeSoto | | 2019-05-24 11:37:38 | View |
Deer slow down litter decomposition by reducing litter quality in a temperate forest
Simon Chollet, Morgane Maillard, Juliane Schorghuber, Sue Grayston, Jean-Louis Martin
https://doi.org/10.1101/690032
Disentangling effects of large herbivores on litter decomposition
Recommended by Sébastien Barot based on reviews by 2 anonymous reviewers
Aboveground – belowground interactions is a fascinating field that has developed in ecology since about 20 years [1]. This field has been very fruitful as measured by the numerous articles published but also by the particular role it has played in the development of soil ecology. While soil ecology has for a long time developed partially independently from “general ecology” [2], the field of aboveground – belowground interactions has shown that all ecological interactions occurring within the soil are likely to impact plant growth and plant physiology because they have their roots within the soil. In turns, this should impact the aerial system of plants (higher or lower biomasses, changes in leaf quality…), which should cascade on the aboveground food web. Conversely, all ecological interactions occurring aboveground likely impact plant growth, which should cascade to their root systems, and thus to the soil functioning and the soil food web (through changes in the emission of exudates or inputs of dead roots…). Basically, plants are linking the belowground and aboveground worlds because, as terrestrial primary producers, they need to have (1) leaves to capture CO2 and exploit light and (2) roots to absorb water and mineral nutrients. The article I presently recommend [3] tackles this general issue through the prism of the impact of large herbivores on the decomposition of leaf litter.
This issue is a relatively old one [4, 5] but still deserves efforts because there have been relatively few studies on the subject and because the issue is relatively complex due to the diversity of mechanisms involved and the difficulty to disentangle them. I recommend this article because the authors have cleverly taken advantage of a ‘‘natural’’ long-term experiment, i.e. three islands with contrasted deer densities, to test whether these large mammals are able to impact leaf litter decomposition and whether they are able to do so through changes in litter quality (because they browse the vegetation) or through changes in soil characteristics (either physical or chemical characteristics or the composition of the decomposer community). They have found that deer decrease litter decomposition, mainly through a decrease in litter quality (increase in its C:N ratio). I particularly appreciate the combination of statistics achieved to test the different hypotheses and the fair and in-depth discussion of the results.
I have to confess that I have two small regrets with this work. First, all replications are implemented within the same three islands, so that it cannot be fully excluded that measured effects should not be attributed to any other possible difference between the three islands. I am fairly sure this is not the case (at least because the three islands have the same environments) but I hope that future studies or meta-analyses will be able analyse independent deer density treatments. Second, as a soil ecologist, I am eager to see results on the decomposer communities, both microorganisms and macrofauna, of the three islands.
References
[1] Hooper, D. U., Bignell, D. E., Brown, V. K., Brussard, L., Dangerfield, J. M., Wall, D. H. and Wolters, V. (2000). Interactions between Aboveground and Belowground Biodiversity in Terrestrial Ecosystems: Patterns, Mechanisms, and Feedbacks. BioScience, 50(12), 1049-1061. doi: 10.1641/0006-3568(2000)050[1049:ibaabb]2.0.co;2
[2] Barot, S., Blouin, M., Fontaine, S., Jouquet, P., Lata, J.-C., and Mathieu, J. (2007). A Tale of Four Stories: Soil Ecology, Theory, Evolution and the Publication System. PLOS ONE, 2(11), e1248. doi: 10.1371/journal.pone.0001248
[3] Chollet S., Maillard M., Schörghuber J., Grayston S. and Martin J.-L. (2019). Deer slow down litter decomposition by reducing litter quality in a temperate forest. bioRxiv, 690032, ver. 3 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/690032
[4] Wardle, D. A., Barker, G. M., Yeates, G. W., Bonner, K. I., and Ghani, A. (2001). Introduced browsing mammals in New Zealand natural forests: aboveground and belowground consequences. Ecological Monographs, 71(4), 587-614. doi: 10.1890/0012-9615(2001)071[0587:ibminz]2.0.co;2
[5] Bardgett, R. D., and Wardle, D. A. (2003). Herbivore-mediated linkages between aboveground and belowground communities. Ecology, 84(9), 2258-2268. doi: 10.1890/02-0274
| Deer slow down litter decomposition by reducing litter quality in a temperate forest | Simon Chollet, Morgane Maillard, Juliane Schorghuber, Sue Grayston, Jean-Louis Martin | <p>In temperate forest ecosystems, the role of deer in litter decomposition, a key nutrient cycling process, remains debated. Deer may modify the decomposition process by affecting plant cover and thus modifying litter abundance. They can also alt... |  | Community ecology, Ecosystem functioning, Herbivory, Soil ecology | Sébastien Barot | | 2019-07-04 14:30:19 | View |
How would variation in environmental predictability affect the use of different learning mechanisms in a social bird?
Recommended by Aliza le Roux based on reviews by Matthew Petelle and 1 anonymous reviewer
In their pre-registered paper [1], McCune and colleagues propose a field-based study of social versus individual learning mechanisms in an avian species (great-tailed grackles) that has been expanding its geographic range. The study forms part of a longer-term project that addresses various aspects of this species’ behaviour and biology, and the experience of the team is clear from the preprint. Assessing variation in learning mechanisms in different sections of the grackles’ distribution range, the researchers will investigate how individual learning and social transmission may impact learning about novel challenges in the environment. Considering that this is a social species, the authors expect both individual learning and social transmission to occur, when groups of grackles encounter new challenges/ opportunities in the wild. This in itself is not a very unusual idea to test [2, 3], but the authors are rigorously distinguishing between imitation, emulation, local enhancement, and social enhancement. Such rigour is certainly valuable in studies of cognition in the wild.
Further, the authors predict that the contribution of individual versus social learning could vary between populations, as the core may contain fewer unfamiliar/novel stimuli than the edge, where artificial sources of water (for example) may be more common. They make an argument that the core, middle, and edge populations would experience differing levels of environmental predictability. If true, their field experiments could yield very novel results on how changes in environmental predictability affect social/individual learning in a single study species. Their data would then give unusual insights into the ecological value of individual learning and distinct forms of social learning – something that is not easy to test in wild animals. The authors consider a variety of alternative hypotheses that may ultimately explain their findings, and clarify their methods and analyses in fine detail. The authors also set out limitations clearly, and give a thorough account of their approaches and thinking.
The reviewers and I have a still-unanswered question, which is central to the study: what is the predictability or unpredictability of the core versus edge environments? Although the authors have explained similarities and distinctions between the different sections of the grackles’ range, their description feels a bit vague -- it's not as rigorous or well-defined as the rest of the paper. Such a lack of definition may be inevitable in the limitations of a preprint, but ultimately it does suggest that there may be real uncertainty about the qualitative differences between the core, edge, and middle environments. The authors do explain that a lack of variation in individual responses to the field experiments would preclude the testing of further hypothesis, but do not mention how a salient lack of variation in novelty/ predictability between the environments could impact their hypotheses.
An assessment/quantification of the rate at which the different populations of grackles encounter novel stimuli would be a cornerstone of the success of this proposed study. Certainly, the authors cannot address this in much more detail during the preprint stage, but they need to consider how to best assess/describe differences before starting the full study. Such an assessment could take the form of either a GIS desktop study (comparing, for example, rates of dam/canal construction in core versus edge sections of the distribution range), or observational/ movement data contrasting how frequently members of core versus edge populations encounter artificial sources of water/food in a given month/year. Considering the long-term nature of the larger project, it is possible that these data are already available, but I am speculating. I would highly recommend that such an assessment be undertaken, beyond the mere mention of expected differences. This would solidify the central idea that there are concrete differences between the environments.
Despite this concern, the authors attended well to the comments and recommendations of the two reviewers – both experts in cognitive ecology. It is a preprint showing clear thinking and a consideration of most of the challenges that may be encountered during the course of the study. My own opinion and the estimations of the two reviewers all underscore the originality and value of this project – this should be a very valuable and potentially novel study. I look forward to seeing the outcomes of the research.
References
[1] McCune, K. B., McElreath, R., and Logan, C. J. (2019). Investigating the use of learning mechanisms in a species that is rapidly expanding its geographic range. In principle recommendation by Peer Community In Ecology. corinalogan.com/Preregistrations/g_sociallearning.html
[2] Benson-Amram, S. and Holekamp, K. E. (2012). Innovative problem solving by wild spotted hyenas. Proceedings of the Royal Society B: Biological Sciences, 279(1744), 4087–4095. doi: 10.1098/rspb.2012.1450
[3] Federspiel, I. G., Boeckle, M., von Bayern, A. M. P. and Emery, N. J. (2019). Exploring individual and social learning in jackdaws (Corvus monedula). Learning & Behavior, 47(3), 258–270. doi: 10.3758/s13420-019-00383-8
| Investigating the use of learning mechanisms in a species that is rapidly expanding its geographic range | Kelsey McCune, Richard McElreath, Corina Logan | This is one of many studies planned for our long-term research on the role of behavior and learning in rapid geographic range expansions. Project background: Behavioral flexibility, the ability to change behavior when circumstances change based on... |  | Behaviour & Ethology, Eco-evolutionary dynamics, Foraging, Preregistrations, Social structure, Spatial ecology, Metacommunities & Metapopulations, Zoology | Aliza le Roux | | 2019-07-23 18:45:20 | View |







