Controversy over the decline of arthropods: a matter of temporal baseline?
François Duchenne, Emmanuelle Porcher, Jean-Baptiste Mihoub, Grégoire Loïs, Colin Fontaine
https://doi.org/10.1101/2022.02.09.479422
Don't jump to conclusions on arthropod abundance dynamics without appropriate data
Recommended by Tim Coulson based on reviews by Gabor L Lovei and 1 anonymous reviewer based on reviews by Gabor L Lovei and 1 anonymous reviewer
Humans are dramatically modifying many aspects of our planet via increasing concentrations of carbon dioxide in the atmosphere, patterns of land-use change, and unsustainable exploitation of the planet’s resources. These changes impact the abundance of species of wild organisms, with winners and losers. Identifying how different species and groups of species are influenced by anthropogenic activity in different biomes, continents, and habitats, has become a pressing scientific question with many publications reporting analyses of disparate data on species population sizes. Many conclusions are based on the linear analysis of rather short time series of organismal abundances.
There has been particular interest in how arthropods are impacted by environmental change, with several recent papers reporting contradictory results. To investigate why these contradictions might arise, Duchenne et al. (2022) conducted an analysis of four published data sets along with a series of experimental analyses of simulated time series to examine the power of widely used statistical analyses to gain inference on temporal trends. Their important paper reveals that accurate inference on dynamics, particularly of species that exhibit large temporal fluctuations in abundance, requires time series that are substantially longer than are typically collected, as well as careful thought as to whether linear models are appropriate. Linear analyses of short time series are susceptible to providing unreliable inference as trends can be strongly influenced by points at either end of the time series.
Duchenne et al.’s paper provides important insight on the conditions when strong inference on temporal trends of arthropod (and other species) abundances can be made, and when they should be treated with caution. They do not doubt that many insect and arachnid species are changing their abundances, and that patterns in these changes may vary spatially. What their results do say is that we should treat grand claims of population recovery or rapid declines apparently to extinction with caution when they are based on short time series, particularly of species that show significant boom and bust dynamics. In many ways, these results are not unexpected, but it is nice to see such careful and thoughtful analyses and interpretation. More data are required for most arthropod species before clear assessments of abundance trends can be made. Given our reliance on many arthropods for food, pollination, and numerous ecosystem services, and the ability of other species to spread devastating human diseases such as dengue and malaria, it is advisable that we slow our modification of their habitats while additional data are collected to allow us to better characterise the trajectory of arthropod populations to understand what the consequences of our actions on the natural world are likely to be.
References
Duchenne F, Porcher E, Mihoub J-B, Loïs G, Fontaine C (2022) Controversy over the decline of arthropods: a matter of temporal baseline? bioRxiv, 2022.02.09.479422, ver. 3 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2022.02.09.479422
| Controversy over the decline of arthropods: a matter of temporal baseline? | François Duchenne, Emmanuelle Porcher, Jean-Baptiste Mihoub, Grégoire Loïs, Colin Fontaine | <p style="text-align: justify;">Recently, a number of studies have reported somewhat contradictory patterns of temporal trends in arthropod abundance, from decline to increase. Arthropods often exhibit non-monotonous variation in abundance over ti... | 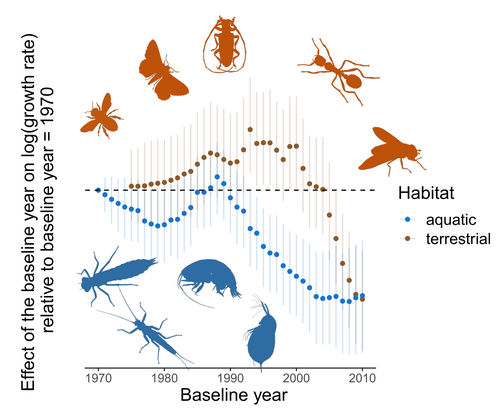 | Conservation biology | Tim Coulson | | 2022-02-11 15:44:44 | View |
Host-mediated, cross-generational intraspecific competition in a herbivore species
Bastien Castagneyrol, Inge van Halder, Yasmine Kadiri, Laura Schillé, Hervé Jactel
https://doi.org/10.1101/2020.07.30.228544
Plants preserve the ghost of competition past for herbivores, but mothers don’t care
Recommended by Sara Magalhães based on reviews by Inês Fragata and Raul Costa-Pereira
Some biological hypotheses are widely popular, so much so that we tend to forget their original lack of success. This is particularly true for hypotheses with catchy names. The ‘Ghost of competition past’ is part of the title of a paper by the great ecologist, JH Connell, one of the many losses of 2020 (Connell 1980). The hypothesis states that, even though we may not detect competition in current populations, their traits and distributions may be shaped by past competition events. Although this hypothesis has known a great success in the ecological literature, the original paper actually ends with “I will no longer be persuaded by such invoking of "the Ghost of Competition Past"”. Similarly, the hypothesis that mothers of herbivores choose host plants where their offspring will have a higher fitness was proposed by John Jaenike in 1978 (Jaenike 1978), and later coined the ‘mother knows best’ hypothesis. The hypothesis was readily questioned or dismissed: “Mother doesn't know best” (Courtney and Kibota 1990), or “Does mother know best?” (Valladares and Lawton 1991), but remains widely popular. It thus seems that catchy names (and the intuitive ideas behind them) have a heuristic value that is independent from the original persuasion in these ideas and the accumulation of evidence that followed it.
The paper by Castagneryol et al. (2021) analyses the preference-performance relationship in the box tree moth (BTM) Cydalima perspectalis, after defoliation of their host plant, the box tree, by conspecifics. It thus has bearings on the two previously mentioned hypotheses. Specifically, they created an artificial population of potted box trees in a greenhouse, in which 60 trees were infested with BTM third instar larvae, whereas 61 were left uninfested. One week later, these larvae were removed and another three weeks later, they released adult BTM females and recorded their host choice by counting egg clutches laid by these females on the plants. Finally, they evaluated the effect of previously infested vs uninfested plants on BTM performance by measuring the weight of third instar larvae that had emerged from those eggs.
This experimental design was adopted because BTM is a multivoltine species. When the second generation of BTM arrives, plants have been defoliated by the first generation and did not fully recover. Indeed, Castagneryol et al. (2021) found that larvae that developed on previously infested plants were much smaller than those developing on uninfested plants, and the same was true for the chrysalis that emerged from those larvae. This provides unequivocal evidence for the existence of a ghost of competition past in this system. However, the existence of this ghost still does not result in a change in the distribution of BTM, precisely because mothers do not know best: they lay as many eggs on plants previously infested than on uninfested plants.
The demonstration that the previous presence of a competitor affects the performance of this herbivore species confirms that ghosts exist. However, whether this entails that previous (interspecific) competition shapes species distributions, as originally meant, remains an open question. Species phenology may play an important role in exposing organisms to the ghost, as this time-lagged competition may have been often overlooked. It is also relevant to try to understand why mothers don’t care in this, and other systems. One possibility is that they will have few opportunities to effectively choose in the real world, due to limited dispersal or to all plants being previously infested.
References
Castagneyrol, B., Halder, I. van, Kadiri, Y., Schillé, L. and Jactel, H. (2021) Host-mediated, cross-generational intraspecific competition in a herbivore species. bioRxiv, 2020.07.30.228544, ver. 5 peer-reviewed and recommended by PCI Ecology. doi: https://doi.org/10.1101/2020.07.30.228544
Connell, J. H. (1980). Diversity and the coevolution of competitors, or the ghost of competition past. Oikos, 131-138. doi: https://doi.org/10.2307/3544421
Courtney, S. P. and Kibota, T. T. (1990) in Insect-plant interactions (ed. Bernays, E.A.) 285-330.
Jaenike, J. (1978). On optimal oviposition behavior in phytophagous insects. Theoretical population biology, 14(3), 350-356. doi: https://doi.org/10.1016/0040-5809(78)90012-6
Valladares, G., and Lawton, J. H. (1991). Host-plant selection in the holly leaf-miner: does mother know best?. The Journal of Animal Ecology, 227-240. doi: https://doi.org/10.2307/5456
| Host-mediated, cross-generational intraspecific competition in a herbivore species | Bastien Castagneyrol, Inge van Halder, Yasmine Kadiri, Laura Schillé, Hervé Jactel | <p>Conspecific insect herbivores co-occurring on the same host plant interact both directly through interference competition and indirectly through exploitative competition, plant-mediated interactions and enemy-mediated interactions. However, the... |  | Competition, Herbivory, Zoology | Sara Magalhães | | 2020-08-03 15:50:23 | View |
Studies of NH4+ and NO3- uptake ability of subalpine plants and resource-use strategy identified by their functional traits
Legay Nicolas, Grassein Fabrice, Arnoldi Cindy, Segura Raphaël, Laîné Philippe, Lavorel Sandra, Clément Jean-Christophe
https://doi.org/10.1101/372235
Nitrate or not nitrate. That is the question
Recommended by Sébastien Barot based on reviews by Vincent Maire and 1 anonymous reviewer
The article by Legay et al. [1] addresses two main issues: the links between belowground and aboveground plant traits and the links between plant strategies (as defined by these traits) and the capacity to absorb nitrate and ammonium. I recommend this work because these are important and current issues. The literature on plant traits is extremely rich and the existence of a leaf economic spectrum linked to a gradient between conservative and acquisitive plants is now extremely well established [2-3]. Many teams are now working on belowground traits and possible links with the aboveground gradients [4-5]. It seems indeed that there is a root economic spectrum but this spectrum is apparently less pronounced than the leaf economic spectrum. The existence of links between the two spectrums are still controversial and are likely not universal as suggested by discrepant results and after all a plant could have a conservative strategy aboveground and an acquisitive strategy belowground (or vice-versa) because, indeed, constraints are different belowground and aboveground (for example because in given ecosystem/vegetation type light may be abundant but not water or mineral nutrients). The various results obtained also suggest that we do not full understand the diversity of belowground strategies, what is at stake with these strategies, and the links with root characteristics.
Each time I give a conference on the work we are carrying out on African grasses that likely absorb ammonium preferentially because they inhibit nitrification [6-7], somebody asks me a question about the fact that plant essentially absorb nitrate because ammonium is toxic and nitrate more available in the soil. The present article confirms that this is not the case and that, though there are currently some teams working on the subject, we do not really know for the moment whether plants absorb nitrate or ammonium, in which proportion, how plastic this proportion is within individuals and within species. This subject seems to me crucial because it is linked to (1) the capacity of ecosystems to conserve nitrogen [8], because nitrate, much more than ammonium, goes out of ecosystems through leaching and denitrification, (2) to carbon cycling and plant energy budget because absorbing nitrate requires spending mucho more energy than absorbing ammonium because nitrate must be reduced before being incorporated in plant biomass, which is very energy costly. These two issues are naturally very relevant to develop efficient cropping systems in terms of carbon and nitrogen.
Interestingly, the present article, comparing three grass species in different sites, suggests that there is no trade-off between the absorption of nitrate and ammonium: more acquisitive individuals tend to absorb more ammonium and nitrate. This is contrary to hypotheses we made to predict the outcome of competition between plants absorbing nitrate and ammonium in different proportions [9] but should be tested in the future comparing many different types of plants. The results also suggest that more conservative plants absorb relatively more ammonium, which makes sense because this allows them to spare the energy necessary to reduce nitrate. This leads to the question of the effect of these strategies on nitrogen retention within the ecosystem. If nitrification is high (low), absorbing ammonium is not efficient and likely leads to high (low) nitrogen losses. This should be tested in the future. Moreover, the authors have measured the absorption of nitrate and ammonium through measurements at the root scale on cut roots. This should be complemented by measurements at the whole plant scale.
References
[1] Legay, N., Grassein, F., Arnoldi, C., Segura, R., Laîné, P., Lavorel, S. and Clément, J.-C. (2020). Studies of NH4+ and NO3- uptake ability of subalpine plants and resource-use strategy identified by their functional traits. bioRxiv, 372235, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/372235
[2] Shipley, B., Lechowicz, M.J., Wright, I. & Reich, P.B. (2006) Fundamental trade-offs generating the worldwide leaf economics spectrum. Ecology, 87, 535-541. doi: 10.1890/05-1051
[3] Reich, P.B. (2014) The world-wide ‘fast-slow’ plant economics spectrum: a traits manifesto. J. Ecol., 102, 275-301. doi: 10.1111/1365-2745.12211
[4] Maire, V., Gross, N., Pontes, L.D.S., Picon-Cochard, C. & Soussana, J.F. (2009) Trade-off between root nitrogen acquisition and shoot nitrogen utilization across 13 co-occurring pasture grass species. Func. Ecol., 23, 668-679. doi: 10.1111/j.1365-2435.2009.01557.x
[5] Roumet, C., Birouste, M., Picon-Cochard, C., Ghestem, M., Osman, N., Vrignon-Brenas, S., Cao, K.F. & Stokes, A. (2016) Root structure-function relationships in 74 species: evidence of a root economics spectrum related to carbon economy. New. Phytol., 210, 815-826. doi: 10.1111/nph.13828
[6] Lata, J.-C., Degrange, V., Raynaud, X., Maron, P.-A., Lensi, R. & Abbadie, L. (2004) Grass populations control nitrification in savanna soils. Funct. Ecol., 18, 605-611. doi: 10.1111/j.0269-8463.2004.00880.x
[7] Srikanthasamy, T., Leloup, J., N’Dri, A.B., Barot, S., Gervaix, J., Koné, A.W., Koffi, K.F., Le Roux, X., Raynaud, X. & Lata, J.-C. (2018) Contrasting effects of grasses and trees on microbial N-cycling in an African humid savanna. Soil Biol. Biochem., 117, 153-163. doi: 10.1016/j.soilbio.2017.11.016
[8] Boudsocq, S., Lata, J.C., Mathieu, J., Abbadie, L. & Barot, S. (2009) Modelling approach to analyze the effects of nitrification inhibition on primary production. Func. Ecol., 23, 220-230. doi: 10.1111/j.1365-2435.2008.01476.x
[9] Boudsocq, S., Niboyet, A., Lata, J.-C., Raynaud, X., Loeuille, N., Mathieu, J., Blouin, M., Abbadie, L. & Barot, S. (2012) Plant preference for ammonium versus nitrate: a neglected determinant of ecosystem functioning? Am. Nat., 180, 60-69. doi: 10.1086/665997
| Studies of NH4+ and NO3- uptake ability of subalpine plants and resource-use strategy identified by their functional traits | Legay Nicolas, Grassein Fabrice, Arnoldi Cindy, Segura Raphaël, Laîné Philippe, Lavorel Sandra, Clément Jean-Christophe | <p>The leaf economics spectrum (LES) is based on a suite of leaf traits related to plant functioning and ranges from resource-conservative to resource-acquisitive strategies. However, the relationships with root traits, and the associated belowgro... |  | Community ecology, Physiology, Terrestrial ecology | Sébastien Barot | | 2018-07-19 14:22:28 | View |
On the efficacy of restoration in stream networks: comments, critiques, and prospective recommendations
David Murray-Stoker
https://doi.org/10.1101/611939
A stronger statistical test of stream restoration experiments
Recommended by Karl Cottenie based on reviews by Eric Harvey and Mariana Perez Rocha
The metacommunity framework acknowledges that local sites are connected to other sites through dispersal, and that these connectivity patterns can influence local dynamics [1]. This framework is slowly moving from a framework that guides fundamental research to being actively applied in for instance a conservation context (e.g. [2]). Swan and Brown [3,4] analyzed the results of a suite of experimental manipulations in headwater and mainstem streams on invertebrate community structure in the context of the metacommunity concept. This was an important contribution to conservation ecology.
However, David Murray-Stoker [5] was not satisfied with their statistical analyses, and recreated, and more importantly, improved their original analyses in the peer-reviewed article. The new analyses are based on a combination of a more consistent site selection, checking the model assumptions, using different estimation procedures, and focusing more on effect size calculations versus statistical significance. This peer-reviewed article is thus the perfect example of the advantages of open research: the original authors making available both the data and their R script files, initially first updating the analyses and results themselves, followed by more in-depth analyses of the original data and question.
This peer reviewed went through a very in-depth process itself, with several rounds of questions and feedback that addressed both the statistical analyses, the interpretation of the results, and the conclusions. It also, however, addressed something that is often harder to provide feedback on, for instance the tone of the argument. I hope that scientists interested in these issues will not only read the final manuscript, but also the different steps of the peer review processes. These are very informative, I think, and provide a more complete picture of mainly the raison for certain decisions.
Not only does this provide the reader interested in stream conservation with the opportunity to make up their own mind on the appropriateness of these decisions, but it could potentially lead to more analyses of this important data set. For instance, maybe a formal meta-analysis that starts with the effect sizes of all the original studies might bring some new insights into this question?
References
[1] Leibold, M. A., Holyoak, M., Mouquet, N. et al. (2004). The metacommunity concept: a framework for multi‐scale community ecology. Ecology letters, 7(7), 601-613. doi: 10.1111/j.1461-0248.2004.00608.x
[2] Heino, J. (2013). The importance of metacommunity ecology for environmental assessment research in the freshwater realm. Biological Reviews, 88(1), 166-178. doi: 10.1111/j.1469-185X.2012.00244.x
[3] Swan, C. M., and Brown, B. L. (2017). Metacommunity theory meets restoration: isolation may mediate how ecological communities respond to stream restoration. Ecological Applications, 27(7), 2209-2219. doi: 10.1002/eap.1602
[4] Swan, C. M., and Brown, B. L. (2018). Erratum for: Metacommunity theory meets restoration: isolation may mediate how ecological communities respond to stream restoration. Ecological Applications 28:1370–1371. doi: 10.1002/eap.1738
[5] Murray-Stoker, D. (2020). On the efficacy of restoration in stream networks: comments, critiques, and prospective recommendations. bioRxiv, 611939, ver. 7 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/611939
| On the efficacy of restoration in stream networks: comments, critiques, and prospective recommendations | David Murray-Stoker | <p>Swan and Brown (2017) recently addressed the effects of restoration on stream communities under the meta-community framework. Using a combination of headwater and mainstem streams, Swan and Brown (2017) evaluated how position within a stream ne... |  | Community ecology, Freshwater ecology, Spatial ecology, Metacommunities & Metapopulations | Karl Cottenie | | 2019-09-21 22:12:57 | View |
A meta-analysis to infer generic predator functional response
Recommended by Samir Simon Suweis based on reviews by Ludek Berec and gyorgy barabas
Species interactions are classically derived from the law of mass action: the probability that, for example, a predation event occurs is proportional to the product of the density of the prey and predator species. In order to describe how predator and prey species populations grow, is then necessary to introduce functional response, describing the intake rate of a consumer as a function of food (e.g. prey) density.
Linear functional responses shapes are typically introduced in the ecological modeling of population dynamics for both predator-prey and mutualistic systems [1,2]. Recently some works have proposed alternatives to the classic approach for mutualistic systems [3,4], both because cooperative interactions also model effect not directly related to mass action [3] and for analytical tractability [4,5].
In this work [6] the authors challenge the classic modeling of functional response also for predator-prey systems. In particular, they use a meta-analysis of several observational studies of predator-prey ecosystems to infer a generic predator functional response, fitting a phenomenological generalization of the mass-action law. Using advanced statistical analysis, they show that the functional response obtained from data is clearly different from the mass-action assumption. In fact, they found that it scales sub-linearly as the square root of the ratio between predator and prey biomass. They further argue that, from a macro-ecological point of view, using such a phenomenological relationship might be more valuable than relying on various mechanistic functional response formulations.
The manuscript thus provides an interesting different perspective on how to approach predator-prey modelling and for this reason, I have recommended the work for PCI Ecology.
References
[1] Volterra, V. (1928). Variations and Fluctuations of the Number of Individuals in Animal Species living together. ICES Journal of Marine Science, 3(1), 3–51. doi: 10.1093/icesjms/3.1.3
[2] Bastolla, U., Fortuna, M. A., Pascual-García, A., Ferrera, A., Luque, B., and Bascompte, J. (2009). The architecture of mutualistic networks minimizes competition and increases biodiversity. Nature, 458(7241), 1018–1020. doi: 10.1038/nature07950
[3] Tu, C., Suweis, S., Grilli, J., Formentin, M., and Maritan, A. (2019). Reconciling cooperation, biodiversity and stability in complex ecological communities. Scientific Reports, 9(1), 1–10. doi: 10.1038/s41598-019-41614-2
[4] García-Algarra, J., Galeano, J., Pastor, J. M., Iriondo, J. M., and Ramasco, J. J. (2014). Rethinking the logistic approach for population dynamics of mutualistic interactions. Journal of Theoretical Biology, 363, 332–343. doi: 10.1016/j.jtbi.2014.08.039
[5] Suweis, S., Simini, F., Banavar, J. R., and Maritan, A. (2013). Emergence of structural and dynamical properties of ecological mutualistic networks. Nature, 500(7463), 449–452. doi: 10.1038/nature12438
[6] Barbier, M., Wojcik, L., and Loreau, M. (2020). A macro-ecological approach to predators’ functional response. BioRxiv, 832220, ver. 4 recommended and peer-reviewed by Peer Community in Ecology. doi: 10.1101/832220
| A macro-ecological approach to predators' functional response | Matthieu Barbier, Laurie Wojcik, Michel Loreau | <p>Predation often deviates from the law of mass action: many micro- and meso-scale experiments have shown that consumption saturates with resource abundance, and decreases due to interference between consumers. But does this observation hold at m... | 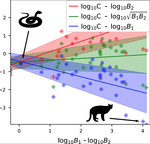 | Community ecology, Food webs, Meta-analyses, Theoretical ecology | Samir Simon Suweis | | 2019-11-08 15:42:16 | View |
The return of the trophic chain: fundamental vs realized interactions in a simple arthropod food web
Inmaculada Torres-Campos, Sara Magalhães, Jordi Moya-Laraño, Marta Montserrat
https://doi.org/10.1101/324178
From deserts to avocado orchards - understanding realized trophic interactions in communities
Recommended by Francis John Burdon based on reviews by Owen Petchey and 2 anonymous reviewers
The late eminent ecologist Gary Polis once stated that “most catalogued food-webs are oversimplified caricatures of actual communities” and are “grossly incomplete representations of communities in terms of both diversity and trophic connections.” Not content with that damning indictment, he went further by railing that “theorists are trying to explain phenomena that do not exist” [1]. The latter critique might have been push back for Robert May´s ground-breaking but ultimately flawed research on the relationship between food-web complexity and stability [2]. Polis was a brilliant ecologist, and his thinking was clearly influenced by his experiences researching desert food webs. Those food webs possess an uncommon combination of properties, such as frequent omnivory, cannibalism, and looping; high linkage density (L/S); and a nearly complete absence of apex consumers, since few species completely lack predators or parasites [3]. During my PhD studies, I was lucky enough to visit Joshua Tree National Park on the way to a conference in New England, and I could immediately see the problems posed by desert ecosystems. At the time, I was ruminating on the “harsh-benign” hypothesis [4], which predicts that the relative importance of abiotic and biotic forces should vary with changes in local environmental conditions (from harsh to benign). Specifically, in more “harsh” environments, abiotic factors should determine community composition whilst weakening the influence of biotic interactions. However, in the harsh desert environment I saw first-hand evidence that species interactions were not diminished; if anything, they were strengthened. Teddy-bear chollas possessed murderously sharp defenses to protect precious water, creosote bushes engaged in belowground “chemical warfare” (allelopathy) to deter potential competitors, and rampant cannibalism amongst scorpions drove temporal and spatial ontogenetic niche partitioning. Life in the desert was hard, but you couldn´t expect your competition to go easy on you.
If that experience colored my thinking about nature’s reaction to a capricious environment, then the seminal work by Robert Paine on the marine rocky shore helped further cement the importance of biotic interactions. The insights provided by Paine [5] brings us closer to the research reported in the preprint “The return of the trophic chain: fundamental vs realized interactions in a simple arthropod food web” [6], given that the authors in that study hold the environment constant and test the interactions between different permutations of a simple community. Paine [5] was able to elegantly demonstrate using the chief protagonist Pisaster ochraceus (a predatory echinoderm also known as the purple sea star) that a keystone consumer could exert strong top-down control that radically reshaped the interactions amongst other community members. What was special about this study was that the presence of Pisaster promoted species diversity by altering competition for space by sedentary species, providing a rare example of an ecological network experiment combining trophic and non-trophic interactions. Whilst there are increasing efforts to describe these interactions (e.g., competition and facilitation) in multiplex networks [7], the authors of “The return of the trophic chain: fundamental vs realized interactions in a simple arthropod food web” [6] have avoided strictly competitive interactions for the sake of simplicity. They do focus on two trophic forms of competition, namely intraguild predation and apparent competition. These two interaction motifs, along with prey switching are relevant to my own research on the influence of cross-ecosystem prey subsidies to receiving food webs [8]. In particular, the apparent competition motif may be particularly important in the context of emergent adult aquatic insects as prey subsidies to terrestrial consumers. This was demonstrated by Henschel et al. [9] where the abundances of emergent adult aquatic midges in riparian fields adjacent to a large river helped stimulate higher abundances of spiders and lower abundances of herbivorous leafhoppers, leading to a trophic cascade. The aquatic insects had a bottom-up effect on spiders and this subsidy facilitated a top-down effect that cascaded from spiders to leafhoppers to plants. The apparent competition motif becomes relevant because the aquatic midges exerted a negative indirect effect on leafhoppers mediated through their common arachnid predators.
In the preprint “The return of the trophic chain: fundamental vs realized interactions in a simple arthropod food web” [6], the authors have described different permutations of a simple mite community present in avocado orchards (Persea americana). This community comprises of two predators (Euseius stipulatus and Neoseiulus californicus), one herbivore as shared prey (Oligonychus perseae), and pollen of Carpobrotus edulis as alternative food resource, with the potential for the intraguild predation and apparent competition interaction motifs to be expressed. The authors determined that these motifs should be realized based off pairwise feeding trials. It is common for food-web researchers to depict potential food webs, which contain all species sampled and all potential trophic links based on laboratory feeding trials (as demonstrated here) or from observational data and literature reviews [10]. In reality, not all these potential feeding links are realized because species may partition space and time, thus driving alternative food-web architectures. In “The return of the trophic chain: fundamental vs realized interactions in a simple arthropod food web” [6], the authors are able to show that placing species in combinations that should yield more complex interaction motifs based off pairwise feeding trials fails to deliver – the predators revert to their preferred prey resulting in modular and simple trophic chains to be expressed. Whilst these realized interaction motifs may be stable, there might also be a tradeoff with function by yielding less top-down control than desirable when considering the potential for ecosystem services such as pest management. These are valuable insights, although it should be noted that here the fundamental niche is described in a strictly Eltonian sense as a trophic role [11]. Adding additional niche dimensions (sensu [12]), such as a thermal gradient could alter the observed interactions, although it might be possible to explain these contingencies through metabolic and optimal foraging theory combined with species traits. Nonetheless, the results of these experiments further demonstrate the need for ecologists to cross-validate theory with empirical approaches to develop more realistic and predictive food-web models, lest they invoke the wrath of Gary Polis´ ghost by “trying to explain phenomena that do not exist”.
References
[1] Polis, G. A. (1991). Complex trophic interactions in deserts: an empirical critique of food-web theory. The American Naturalist, 138(1), 123-155. doi: 10.1086/285208
[2] May, R. M. (1973). Stability and complexity in model ecosystems. Princeton University Press, Princeton, NJ, USA
[3] Dunne, J. A. (2006). The network structure of food webs. In Pascual, M., & Dunne, J. A. (eds) Ecological Networks: Linking Structure to Dynamics in Food Webs. Oxford University Press, New York, USA, 27-86
[4] Menge, B. A., & Sutherland, J. P. (1976). Species diversity gradients: synthesis of the roles of predation, competition, and temporal heterogeneity. The American Naturalist, 110(973), 351-369. doi: 10.1086/283073
[5] Paine, R. T. (1966). Food web complexity and species diversity. The American Naturalist, 100(910), 65-75. doi: 10.1086/282400
[6] Torres-Campos, I., Magalhães, S., Moya-Laraño, J., & Montserrat, M. (2018). The return of the trophic chain: fundamental vs realized interactions in a simple arthropod food web. bioRxiv, 324178, ver. 5 peer-reviewed and recommended by PCI Ecol. doi: 10.1101/324178
[7] Kéfi, S., Berlow, E. L., Wieters, E. A., Joppa, L. N., Wood, S. A., Brose, U., & Navarrete, S. A. (2015). Network structure beyond food webs: mapping non‐trophic and trophic interactions on Chilean rocky shores. Ecology, 96(1), 291-303. doi: 10.1890/13-1424.1
[8] Burdon, F. J., & Harding, J. S. (2008). The linkage between riparian predators and aquatic insects across a stream‐resource spectrum. Freshwater Biology, 53(2), 330-346. doi: 10.1111/j.1365-2427.2007.01897.x
[9] Henschel, J. R., Mahsberg, D., & Stumpf, H. (2001). Allochthonous aquatic insects increase predation and decrease herbivory in river shore food webs. Oikos, 93(3), 429-438. doi: 10.1034/j.1600-0706.2001.930308.x
[10] Brose, U., Pavao-Zuckerman, M., Eklöf, A., Bengtsson, J., Berg, M. P., Cousins, S. H., Mulder, C., Verhoef, H. A., & Wolters, V. (2005). Spatial aspects of food webs. In de Ruiter, P., Wolters, V., Moore, J. C., & Melville-Smith, K. (eds) Dynamic Food Webs. vol 3. Academic Press, Burlington, 463-469
[11] Elton, C. (1927). Animal Ecology. Sidgwick and Jackson, London, UK
[12] Hutchinson, G. E. (1957). Concluding Remarks. Cold Spring Harbor Symposia on Quantitative Biology, 22, 415-427. doi: 10.1101/sqb.1957.022.01.039
| The return of the trophic chain: fundamental vs realized interactions in a simple arthropod food web | Inmaculada Torres-Campos, Sara Magalhães, Jordi Moya-Laraño, Marta Montserrat | <p>The mathematical theory describing small assemblages of interacting species (community modules or motifs) has proved to be essential in understanding the emergent properties of ecological communities. These models use differential equations to ... |  | Community ecology, Experimental ecology | Francis John Burdon | | 2018-05-16 19:34:10 | View |
Uncertain predictions of species responses to perturbations lead to underestimate changes at ecosystem level in diverse systems
Recommended by Elisa Thebault based on reviews by Carlos Melian and 1 anonymous reviewer
Different sources of uncertainty are known to affect our ability to predict ecological dynamics (Petchey et al. 2015). However, the consequences of uncertainty on prediction biases have been less investigated, especially when predictions are scaled up to higher levels of organisation as is commonly done in ecology for instance. The study of Orr et al. (2020) addresses this issue. It shows that, in complex systems, the uncertainty of unbiased predictions at a lower level of organisation (e.g. species level) leads to a bias towards underestimation of change at higher level of organisation (e.g. ecosystem level). This bias is strengthened by larger uncertainty and by higher dimensionality of the system.
This general result has large implications for many fields of science, from economics to energy supply or demography. In ecology, as discussed in this study, these results imply that the uncertainty of predictions of species’ change increases the probability of underestimation of changes of diversity and stability at community and ecosystem levels, especially when species richness is high. The uncertainty of predictions of species’ change also increases the probability of underestimation of change when multiple ecosystem functions are considered at once, or when the combined effect of multiple stressors is considered.
The consequences of species diversity on ecosystem functions and stability have received considerable attention during the last decades (e.g. Cardinale et al. 2012, Kéfi et al. 2019). However, since the bias towards underestimation of change increases with species diversity, future studies will need to investigate how the general statistical effect outlined by Orr et al. might affect our understanding of the well-known relationships between species diversity and ecosystem functioning and stability in response to perturbations.
References
Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, Venail P, Narwani A, Mace GM, Tilman D, Wardle DA, Kinzig AP, Daily GC, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S (2012) Biodiversity loss and its impact on humanity. Nature, 486, 59–67. https://doi.org/10.1038/nature11148
Kéfi S, Domínguez‐García V, Donohue I, Fontaine C, Thébault E, Dakos V (2019) Advancing our understanding of ecological stability. Ecology Letters, 22, 1349–1356. https://doi.org/10.1111/ele.13340
Orr JA, Piggott JJ, Jackson A, Arnoldi J-F (2020) Why scaling up uncertain predictions to higher levels of organisation will underestimate change. bioRxiv, 2020.05.26.117200. https://doi.org/10.1101/2020.05.26.117200
Petchey OL, Pontarp M, Massie TM, Kéfi S, Ozgul A, Weilenmann M, Palamara GM, Altermatt F, Matthews B, Levine JM, Childs DZ, McGill BJ, Schaepman ME, Schmid B, Spaak P, Beckerman AP, Pennekamp F, Pearse IS (2015) The ecological forecast horizon, and examples of its uses and determinants. Ecology Letters, 18, 597–611. https://doi.org/10.1111/ele.12443
| Why scaling up uncertain predictions to higher levels of organisation will underestimate change | James Orr, Jeremy Piggott, Andrew Jackson, Jean-François Arnoldi | <p>Uncertainty is an irreducible part of predictive science, causing us to over- or underestimate the magnitude of change that a system of interest will face. In a reductionist approach, we may use predictions at the level of individual system com... |  | Community ecology, Ecosystem functioning, Theoretical ecology | Elisa Thebault | Anonymous | 2020-06-02 15:41:12 | View |
Deer slow down litter decomposition by reducing litter quality in a temperate forest
Simon Chollet, Morgane Maillard, Juliane Schorghuber, Sue Grayston, Jean-Louis Martin
https://doi.org/10.1101/690032
Disentangling effects of large herbivores on litter decomposition
Recommended by Sébastien Barot based on reviews by 2 anonymous reviewers
Aboveground – belowground interactions is a fascinating field that has developed in ecology since about 20 years [1]. This field has been very fruitful as measured by the numerous articles published but also by the particular role it has played in the development of soil ecology. While soil ecology has for a long time developed partially independently from “general ecology” [2], the field of aboveground – belowground interactions has shown that all ecological interactions occurring within the soil are likely to impact plant growth and plant physiology because they have their roots within the soil. In turns, this should impact the aerial system of plants (higher or lower biomasses, changes in leaf quality…), which should cascade on the aboveground food web. Conversely, all ecological interactions occurring aboveground likely impact plant growth, which should cascade to their root systems, and thus to the soil functioning and the soil food web (through changes in the emission of exudates or inputs of dead roots…). Basically, plants are linking the belowground and aboveground worlds because, as terrestrial primary producers, they need to have (1) leaves to capture CO2 and exploit light and (2) roots to absorb water and mineral nutrients. The article I presently recommend [3] tackles this general issue through the prism of the impact of large herbivores on the decomposition of leaf litter.
This issue is a relatively old one [4, 5] but still deserves efforts because there have been relatively few studies on the subject and because the issue is relatively complex due to the diversity of mechanisms involved and the difficulty to disentangle them. I recommend this article because the authors have cleverly taken advantage of a ‘‘natural’’ long-term experiment, i.e. three islands with contrasted deer densities, to test whether these large mammals are able to impact leaf litter decomposition and whether they are able to do so through changes in litter quality (because they browse the vegetation) or through changes in soil characteristics (either physical or chemical characteristics or the composition of the decomposer community). They have found that deer decrease litter decomposition, mainly through a decrease in litter quality (increase in its C:N ratio). I particularly appreciate the combination of statistics achieved to test the different hypotheses and the fair and in-depth discussion of the results.
I have to confess that I have two small regrets with this work. First, all replications are implemented within the same three islands, so that it cannot be fully excluded that measured effects should not be attributed to any other possible difference between the three islands. I am fairly sure this is not the case (at least because the three islands have the same environments) but I hope that future studies or meta-analyses will be able analyse independent deer density treatments. Second, as a soil ecologist, I am eager to see results on the decomposer communities, both microorganisms and macrofauna, of the three islands.
References
[1] Hooper, D. U., Bignell, D. E., Brown, V. K., Brussard, L., Dangerfield, J. M., Wall, D. H. and Wolters, V. (2000). Interactions between Aboveground and Belowground Biodiversity in Terrestrial Ecosystems: Patterns, Mechanisms, and Feedbacks. BioScience, 50(12), 1049-1061. doi: 10.1641/0006-3568(2000)050[1049:ibaabb]2.0.co;2
[2] Barot, S., Blouin, M., Fontaine, S., Jouquet, P., Lata, J.-C., and Mathieu, J. (2007). A Tale of Four Stories: Soil Ecology, Theory, Evolution and the Publication System. PLOS ONE, 2(11), e1248. doi: 10.1371/journal.pone.0001248
[3] Chollet S., Maillard M., Schörghuber J., Grayston S. and Martin J.-L. (2019). Deer slow down litter decomposition by reducing litter quality in a temperate forest. bioRxiv, 690032, ver. 3 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/690032
[4] Wardle, D. A., Barker, G. M., Yeates, G. W., Bonner, K. I., and Ghani, A. (2001). Introduced browsing mammals in New Zealand natural forests: aboveground and belowground consequences. Ecological Monographs, 71(4), 587-614. doi: 10.1890/0012-9615(2001)071[0587:ibminz]2.0.co;2
[5] Bardgett, R. D., and Wardle, D. A. (2003). Herbivore-mediated linkages between aboveground and belowground communities. Ecology, 84(9), 2258-2268. doi: 10.1890/02-0274
| Deer slow down litter decomposition by reducing litter quality in a temperate forest | Simon Chollet, Morgane Maillard, Juliane Schorghuber, Sue Grayston, Jean-Louis Martin | <p>In temperate forest ecosystems, the role of deer in litter decomposition, a key nutrient cycling process, remains debated. Deer may modify the decomposition process by affecting plant cover and thus modifying litter abundance. They can also alt... |  | Community ecology, Ecosystem functioning, Herbivory, Soil ecology | Sébastien Barot | | 2019-07-04 14:30:19 | View |
Diagnosis of planktonic trophic network dynamics with sharp qualitative changes
Cedric Gaucherel, Stolian Fayolle, Raphael Savelli, Olivier Philippine, Franck Pommereau, Christine Dupuy
https://doi.org/10.1101/2023.06.29.547055
A new approach to describe qualitative changes of complex trophic networks
Recommended by Francis Raoul based on reviews by Tim Coulson and 1 anonymous reviewer
Modelling the temporal dynamics of trophic networks has been a key challenge for community ecologists for decades, especially when anthropogenic and natural forces drive changes in species composition, abundance, and interactions over time. So far, most modelling methods fail to incorporate the inherent complexity of such systems, and its variability, to adequately describe and predict temporal changes in the topology of trophic networks.
Taking benefit from theoretical computer science advances, Gaucherel and colleagues (2024) propose a new methodological framework to tackle this challenge based on discrete-event Petri net methodology. To introduce the concept to naïve readers the authors provide a useful example using a simplistic predator-prey model.
The core biological system of the article is a freshwater trophic network of western France in the Charente-Maritime marshes of the French Atlantic coast. A directed graph describing this system was constructed to incorporate different functional groups (phytoplankton, zooplankton, resources, microbes, and abiotic components of the environment) and their interactions. Rules and constraints were then defined to, respectively, represent physiochemical, biological, or ecological processes linking network components, and prevent the model from simulating unrealistic trajectories. Then the full range of possible trajectories of this mechanistic and qualitative model was computed.
The model performed well enough to successfully predict a theoretical trajectory plus two trajectories of the trophic network observed in the field at two different stations, therefore validating the new methodology introduced here. The authors conclude their paper by presenting the power and drawbacks of such a new approach to qualitatively model trophic networks dynamics.
Reference
Cedric Gaucherel, Stolian Fayolle, Raphael Savelli, Olivier Philippine, Franck Pommereau, Christine Dupuy (2024) Diagnosis of planktonic trophic network dynamics with sharp qualitative changes. bioRxiv 2023.06.29.547055, ver. 2 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2023.06.29.547055
| Diagnosis of planktonic trophic network dynamics with sharp qualitative changes | Cedric Gaucherel, Stolian Fayolle, Raphael Savelli, Olivier Philippine, Franck Pommereau, Christine Dupuy | <p>Trophic interaction networks are notoriously difficult to understand and to diagnose (i.e., to identify contrasted network functioning regimes). Such ecological networks have many direct and indirect connections between species, and these conne... | 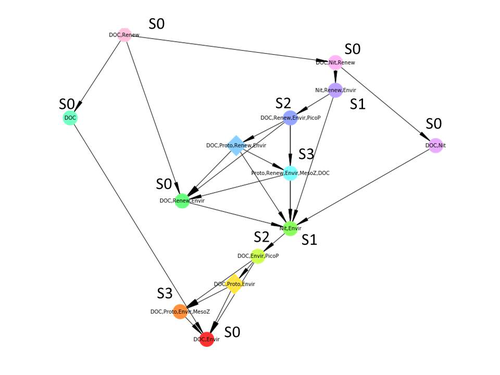 | Community ecology, Ecosystem functioning, Food webs, Freshwater ecology, Interaction networks, Microbial ecology & microbiology | Francis Raoul | Tim Coulson | 2023-07-03 10:42:34 | View |
Intraspecific diversity loss in a predator species alters prey community structure and ecosystem functions
Allan Raffard, Julien Cucherousset, José M. Montoya, Murielle Richard, Samson Acoca-Pidolle, Camille Poésy, Alexandre Garreau, Frédéric Santoul & Simon Blanchet.
https://doi.org/10.1101/2020.06.10.144337
Hidden diversity: how genetic richness affects species diversity and ecosystem processes in freshwater ponds
Recommended by Frederik De Laender based on reviews by Andrew Barnes and Jes Hines
Biodiversity loss can have important consequences for ecosystem functions, as exemplified by a large body of literature spanning at least three decades [1–3]. While connections between species diversity and ecosystem functions are now well-defined and understood, the importance of diversity within species is more elusive. Despite a surge in theoretical work on how intraspecific diversity can affect coexistence in simple community types [4,5], not much is known about how intraspecific diversity drives ecosystem processes in more complex community types. One particular challenge is that intraspecific diversity can be expressed as observable variation of functional traits, or instead subsist as genetic variation of which the consequences for ecosystem processes are harder to grasp.
Raffard et al. [6] examined how intraspecific biodiversity loss in a consumer fish changes species diversity at lower trophic levels and ecosystem processes in pond mesocosms. An interesting feature of this experiment is that it crosses functional and genetic intraspecific diversity. To do so, Raffard and colleagues measured and genotyped European minnow (P. phoxinus) individuals sampled from streams across southern France. Combining these collected specimens into experimental ponds allowed them to control functional (population variance of body size) and genetic intraspecific richness (number of genotypes).
Effects on minnow biomass production were mostly small; biomass was significantly reduced only when lowering both functional and genetic richness. However, the consequences for lower trophic levels (zooplankton and macroinvertebrates) were more pronounced and – importantly – not intuitive. For instance, the macroinvertebrate community was less species-diverse at higher minnow functional richness. If minnows with different body sizes would be the main regulator factors [7] explaining macroinvertebrate interactions, one would expect a more diverse set of minnow body sizes (i.e. higher functional minnow richness) to permit higher instead of lower macroinvertebrate richness. At the same time, the macroinvertebrate community was more species-diverse at higher minnow genotype richness, which could indicate unobserved minnow traits determining macroinvertebrate diversity more than the usual suspects (functional consumer richness). Such unobserved traits could be behavioral traits, allowing for resource partitioning among fish.
The consequences of functional minnow diversity loss on zooplankton diversity were negative, as expected in case body size differences among fish would facilitate coexistence of their zooplankton prey, as explained above. However, this was only the case when genetic diversity was high, suggesting nonstraightforward interactive effects of observed and non-observed traits on prey diversity.
The effects of functional and genetic minnow diversity loss on invertebrate (macroinvertebrates and zooplankton) abundance were more consistent than for invertebrate diversity. This suggests again nonstraightforward relationships in this experimental ecosystem, but now between invertebrate diversity and abundance. When using abundance as a proxy for an ecosystem process (which the authors did not), this result illustrates that biodiversity loss in multitrophic communities can have consequences that are challenging to interpret, let alone predict [8,9]. Path analyses showed how the observed changes of invertebrate diversity and abundance co-determined decomposition, a key ecosystem function. These path analyses had highest explanatory power show when including both kinds of intraspecific diversity.
Taken together, the results by Raffard and colleagues suggest that genetic consumer richness can drive species diversity of connected trophic levels and ecosystem processes with similar magnitude as functional diversity. Indeed, the effects of genetic consumer richness were shown to be so strong as to compensate or exacerbate the loss of observed functional richness. The exact mechanisms explaining these effects remain to be identified, however. The possibility that fish grazing by fish with different (observed or not observed) traits regulates coexistence among invertebrate prey, for instance, would depend on how strong fish consumption feeds back on prey growth during a 30-week experiment. As the authors indicate, detailed studies on resource partitioning among consumers (e.g. using stable isotope labelling) can shed light on these matters. Doing so may address a more fundamental question, which is if the mechanisms linking intraspecific diversity to function are different from those linking interspecific diversity to function, and at what time scales.
References
[1] Tilman D, Downing JA (1994) Biodiversity and stability in grasslands. Nature, 367, 363–365. https://doi.org/10.1038/367363a0
[2] Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, Venail P, Narwani A, Mace GM, Tilman D, Wardle DA, Kinzig AP, Daily GC, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S (2012) Biodiversity loss and its impact on humanity. Nature, 486, 59–67. https://doi.org/10.1038/nature11148
[3] De Laender F, Rohr JR, Ashauer R, Baird DJ, Berger U, Eisenhauer N, Grimm V, Hommen U, Maltby L, Meliàn CJ, Pomati F, Roessink I, Radchuk V, Brink PJV den (2016) Reintroducing Environmental Change Drivers in Biodiversity–Ecosystem Functioning Research. Trends in Ecology & Evolution, 31, 905–915. https://doi.org/10.1016/j.tree.2016.09.007
[4] Hart SP, Schreiber SJ, Levine JM (2016) How variation between individuals affects species coexistence. Ecology Letters, 19, 825–838. https://doi.org/10.1111/ele.12618
[5] Barabás G, D’Andrea R (2016) The effect of intraspecific variation and heritability on community pattern and robustness. Ecology Letters, 19, 977–986. https://doi.org/10.1111/ele.12636
[6] Raffard A, Cucherousset J, Montoya JM, Richard M, Acoca-Pidolle S, Poésy C, Garreau A, Santoul F, Blanchet S (2020) Intraspecific diversity loss in a predator species alters prey community structure and ecosystem functions. bioRxiv, 2020.06.10.144337, ver. 3 peer-reviewed and recommended by PCI Ecology. https://doi.org/10.1101/2020.06.10.144337
[7] Pásztor L, Botta-Dukát Z, Magyar G, Czárán T, Meszéna G. Theory-Based Ecology: A Darwinian approach. Oxford University Press. https://doi.org/10.1093/acprof:oso/9780199577859.001.0001
[8] Binzer A, Guill C, Rall BC, Brose U (2016) Interactive effects of warming, eutrophication and size structure: impacts on biodiversity and food-web structure. Global Change Biology, 22, 220–227. https://doi.org/10.1111/gcb.13086
[9] Schwarz B, Barnes AD, Thakur MP, Brose U, Ciobanu M, Reich PB, Rich RL, Rosenbaum B, Stefanski A, Eisenhauer N (2017) Warming alters energetic structure and function but not resilience of soil food webs. Nature Climate Change, 7, 895–900. https://doi.org/10.1038/s41558-017-0002-z
| Intraspecific diversity loss in a predator species alters prey community structure and ecosystem functions | Allan Raffard, Julien Cucherousset, José M. Montoya, Murielle Richard, Samson Acoca-Pidolle, Camille Poésy, Alexandre Garreau, Frédéric Santoul & Simon Blanchet. | <p>Loss in intraspecific diversity can alter ecosystem functions, but the underlying mechanisms are still elusive, and intraspecific biodiversity-ecosystem function relationships (iBEF) have been restrained to primary producers. Here, we manipulat... | 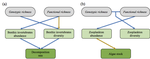 | Community ecology, Ecosystem functioning, Experimental ecology, Food webs, Freshwater ecology | Frederik De Laender | Andrew Barnes | 2020-06-15 09:04:53 | View |







