Understanding geographic range expansions in human-dominated landscapes: does behavioral flexibility modulate flexibility in foraging and social behavior?
Recommended by Julia Astegiano and Esther Sebastián González based on reviews by Pizza Ka Yee Chow and Esther Sebastián González and Esther Sebastián González based on reviews by Pizza Ka Yee Chow and Esther Sebastián González
Which biological traits modulate species distribution has historically been and still is one of the core questions of the macroecology and biogeography agenda [1, 2]. As most of the Earth surface has been modified by human activities [3] understanding the strategies that allow species to inhabit human-dominated landscapes will be key to explain species geographic distribution in the Anthropocene. In this vein, Logan et al. [4] are working on a long-term and integrative project aimed to investigate how great-tailed grackles rapidly expanded their geographic range into North America [4]. Particularly, they want to determine which is the role of behavioral flexibility, i.e. an individual’s ability to modify its behavior when circumstances change based on learning from previous experience [5], in rapid geographic range expansions. The authors are already working in a set of complementary questions described in pre-registrations that have already been recommended at PCI Ecology: (1) Do individuals with greater behavioral flexibility rely more on causal cognition [6]? (2) Which are the mechanisms that lead to behavioral flexibility [7]? (3) Does the manipulation of behavioral flexibility affect exploration, but not boldness, persistence, or motor diversity [8]? (4) Can context changes improve behavioral flexibility [9]?
In this new pre-registration, they aim to determine whether the more behaviorally flexible individuals have more flexible foraging behaviors (i.e. use a wider variety of foraging techniques in the wild and eat a larger number of different foods), habitat use (i.e. higher microhabitat richness) and social relationships (i.e., are more likely to have a greater number of bonds or stronger bonds with other individuals; [4]). The project is ambitious, combining both the experimental characterization of individuals’ behavioral flexibility and the field characterization of the foraging and social behavior of those individuals and of wild ones.
The current great-tailed grackles project will be highly relevant to understand rapid geographic range expansions in a changing world. In this vein, this pre-registration will particularly help to go one step further in our understanding of behavioral flexibility as a determinant of species geographic distribution. Logan et al. [4] pre-registration is very well designed, main and alternative hypotheses have been thought and written and methods are presented in a very detailed way, which includes the R codes that authors will use in their analyses. Authors have answered in a very detailed way each comment that reviewers have pointed out and modified the pre-registration accordingly, which we consider highly improved the quality of this work. That is why we strongly recommend this pre-registration and look forward to see the results.
References
[1] Gaston K. J. (2003) The structure and dynamics of geographic ranges. Oxford series in Ecology and Evolution. Oxford University Press, New York.
[2] Castro-Insua, A., Gómez‐Rodríguez, C., Svenning, J.C., and Baselga, A. (2018) A new macroecological pattern: The latitudinal gradient in species range shape. Global ecology and biogeography, 27(3), 357-367. doi: 10.1111/geb.12702
[3] Newbold, T., Hudson, L. N., Hill, S. L. L., Contu, S., Lysenko, I., Senior, R. A., et al. (2015). Global effects of land use on local terrestrial biodiversity. Nature, 520(7545), 45–50. doi: 10.1038/nature14324
[4] Logan CJ, McCune K, Bergeron L, Folsom M, Lukas D. (2019). Is behavioral flexibility related to foraging and social behavior in a rapidly expanding species? In principle recommendation by Peer Community In Ecology. http://corinalogan.com/Preregistrations/g_flexforaging.html
[5] Mikhalevich, I., Powell, R., and Logan, C. (2017). Is Behavioural Flexibility Evidence of Cognitive Complexity? How Evolution Can Inform Comparative Cognition. Interface Focus 7: 20160121. doi: 10.1098/rsfs.2016.0121.
[6] Fronhofer, E. (2019) From cognition to range dynamics: advancing our understanding of macroecological patterns. Peer Community in Ecology, 100014. doi: 10.24072/pci.ecology.100014
[7] Vogel, E. (2019) Adapting to a changing environment: advancing our understanding of the mechanisms that lead to behavioral flexibility. Peer Community in Ecology, 100016. doi: 10.24072/pci.ecology.100016
[8] Van Cleve, J. (2019) Probing behaviors correlated with behavioral flexibility. Peer Community in Ecology, 100020. doi: 10.24072/pci.ecology.100020
[9] Coulon, A. (2019) Can context changes improve behavioral flexibility? Towards a better understanding of species adaptability to environmental changes. Peer Community in Ecology, 100019. doi: 10.24072/pci.ecology.100019
| Is behavioral flexibility related to foraging and social behavior in a rapidly expanding species? | Corina Logan, Luisa Bergeron, Carolyn Rowney, Kelsey McCune, Dieter Lukas | This is one of the first studies planned for our long-term research on the role of behavioral flexibility in rapid geographic range expansions. Project background: Behavioral flexibility, the ability to change behavior when circumstances change ba... |  | Behaviour & Ethology, Preregistrations, Zoology | Julia Astegiano | | 2018-10-23 00:47:03 | View |
Adapting to a changing environment: advancing our understanding of the mechanisms that lead to behavioral flexibility
Recommended by Erin Vogel based on reviews by Simon Gingins and 2 anonymous reviewers
Behavioral flexibility is essential for organisms to adapt to an ever-changing environment. However, the mechanisms that lead to behavioral flexibility and understanding what traits makes a species better able to adapt behavior to new environments has been understudied. Logan and colleagues have proposed to use a series of experiments, using great-tailed grackles as a study species, to test four main hypotheses. These hypotheses are centered around exploring the relationship between behavioral flexibility and inhibition in grackles. This current preregistration is a part of a larger integrative research plan examining behavioral flexibility when faced with environmental change. In this part of the project they will examine specifically if individuals that are more flexible are also better at inhibiting: in other words: they will test the assumption that inhibition is required for flexibility.
First, they will test the hypothesis that behavioral flexibility is manipulatable by using a serial reversal learning task. Second, they will test the hypothesis that manipulating behavioral flexibility (improving reversal learning speed through serial reversals using colored tubers) improves flexibility (rule switching) and problem solving in a new context (multi‑access box and serial reversals on a touch screen). Third, they will test the hypothesis that behavioral flexibility within a context is repeatable within individuals, which is important to test if performance is state dependent. Finally, they will test a fourth hypothesis that individuals should converge on an epsilon‑first learning strategy (learn the correct choice after one trial) as they progress through serial reversals. Their innovative approach using three main tasks (delay of gratification, go-no, detour) will allow them to assess different aspects of inhibitory control. They will analyze the results of all three experiments to also assess the utility of these experiments for studying the potential relationship between inhibition and behavioral flexibility.
In their preregistration, Logan and colleagues have proposed to test these hypotheses, each with a set of testable predictions that can be examined with detailed and justified methodologies. They have also provided a comprehensive plan for analyzing the data. All of the reviewers and I agree that this is a very interesting study that has the potential to answer important questions about a critical topic in behavioral ecology: the role of inhibition in the evolution of behavioral flexibility. Given the positive reviews, the comprehensive responses by the PI and her colleagues, and careful revisions, I highly recommend this preregistration.
| Are the more flexible great-tailed grackles also better at inhibition? | Corina Logan, Kelsey McCune, Zoe Johnson-Ulrich, Luisa Bergeron, Carolyn Rowney, Benjamin Seitz, Aaron Blaisdell, Claudia Wascher | This is a PREREGISTRATION. The DOI was issued by OSF and refers to the whole GitHub repository, which contains multiple files. The specific file we are submitting is g_inhibition.Rmd, which is easily accessible at GitHub at https://github.com/cori... |  | Behaviour & Ethology, Preregistrations, Zoology | Erin Vogel | | 2018-10-12 18:36:00 | View |
Is behavioral flexibility manipulatable and, if so, does it improve flexibility and problem solving in a new context?
Corina Logan, Carolyn Rowney, Luisa Bergeron, Benjamin Seitz, Aaron Blaisdell, Zoe Johnson-Ulrich, Kelsey McCune
http://corinalogan.com/Preregistrations/g_flexmanip.html
Can context changes improve behavioral flexibility? Towards a better understanding of species adaptability to environmental changes
Recommended by Aurélie Coulon based on reviews by Maxime Dahirel and Andrea Griffin
Behavioral flexibility is a key for species adaptation to new environments. Predicting species responses to new contexts hence requires knowledge on the amount to and conditions in which behavior can be flexible. This is what Logan and collaborators propose to assess in a series of experiments on the great-tailed grackles, in a context of rapid range expansion. This pre-registration is integrated into this large research project and concerns more specifically the manipulability of the cognitive aspects of behavioral flexibility. Logan and collaborators will use reversal learning tests to test whether (i) behavioral flexibility is manipulatable, (ii) manipulating flexibility improves flexibility and problem solving in a new context, (iii) flexibility is repeatable within individuals, (iv) individuals are faster at problem solving as they progress through serial reversals. The pre-registration carefully details the hypotheses, their associated predictions and alternatives, and the plan of statistical analyses, including power tests. The ambitious program presented in this pre-registration has the potential to provide important pieces to better understand the mechanisms of species adaptability to new environments.
| Is behavioral flexibility manipulatable and, if so, does it improve flexibility and problem solving in a new context? | Corina Logan, Carolyn Rowney, Luisa Bergeron, Benjamin Seitz, Aaron Blaisdell, Zoe Johnson-Ulrich, Kelsey McCune | This is one of the first studies planned for our long-term research on the role of behavioral flexibility in rapid geographic range expansions. Behavioral flexibility, the ability to adapt behavior to new circumstances, is thought to play an impor... |  | Behaviour & Ethology, Preregistrations, Zoology | Aurélie Coulon | | 2018-07-03 13:23:10 | View |
Heather pollen is not necessarily a healthy diet for bumble bees
Clément Tourbez, Irène Semay, Apolline Michel, Denis Michez, Pascal Gerbaux, Antoine Gekière, Maryse Vanderplanck
https://doi.org/10.5281/zenodo.8192036
The importance of understanding bee nutrition
Recommended by Ignasi Bartomeus based on reviews by Cristina Botías and 1 anonymous reviewer
Contrasting with the great alarm on bee declines, it is astonishing how little basic biology we know about bees, including on abundant and widespread species that are becoming model species. Plant-pollinator relationships are one of the cornerstones of bee ecology, and researchers are increasingly documenting bees' diets. However, we rarely know which effects feeding on different flowers has on bees' health. This paper (Tourbez et al. 2023) uses an elegant experimental setting to test the effect of heather pollen on bumblebees' (Bombus terrestris) reproductive success. This is a timely question as heather is frequently used by bumblebees, and its nectar has been reported to reduce parasite infections. In fact, it has been suggested that bumblebees can medicate themselves when infected (Richardson et al. 2014), and the pollen of some Asteraceae has been shown to help them fight parasites (Gekière et al. 2022). The starting hypothesis is that heather pollen contains flavonoids that might have a similar effect. Unfortunately, Tourbez and collaborators do not support this hypothesis, showing a negative effect of heather pollen, in particular its flavonoids, in bumblebees offspring, and an increase in parasite loads when fed on flavonoids. This is important because it challenges the idea that many pollen and nectar chemical compounds might have a medicinal use, and force us to critically analyze the effect of chemical compounds in each particular case. The results open several questions, such as why bumblebees collect heather pollen, or in which concentrations or pollen mixes it is deleterious. A limitation of the study is that it uses micro-colonies, and extrapolating this to real-world conditions is always complex. Understanding bee declines require a holistic approach starting with bee physiology and scaling up to multispecies population dynamics.
References
Gekière, A., Semay, I., Gérard, M., Michez, D., Gerbaux, P., & Vanderplanck, M. 2022. Poison or Potion: Effects of Sunflower Phenolamides on Bumble Bees and Their Gut Parasite. Biology, 11(4), 545. https://doi.org/10.3390/biology11040545
Richardson, L.L., Adler, L.S., Leonard, A.S., Andicoechea, J., Regan, K.H., Anthony, W.E., Manson, J.S., & Irwin, R.E. 2015. Secondary metabolites in floral nectar reduce parasite infections in bumblebees. Proceedings of the Royal Society of London B: Biological Sciences 282 (1803), 20142471. https://doi.org/10.1098/rspb.2014.2471
Tourbez, C., Semay, I., Michel, A., Michez, D., Gerbaux, P., Gekière A. & Vanderplanck, M. 2023. Heather pollen is not necessarily a healthy diet for bumble bees. Zenodo, ver 3, reviewed and recommended by PCI Ecology. https://doi.org/10.5281/zenodo.8192036
| Heather pollen is not necessarily a healthy diet for bumble bees | Clément Tourbez, Irène Semay, Apolline Michel, Denis Michez, Pascal Gerbaux, Antoine Gekière, Maryse Vanderplanck | <p>There is evidence that specialised metabolites of flowering plants occur in both vegetative parts and floral resources (i.e., pollen and nectar), exposing pollinators to their biological activities. While such metabolites may be toxic to bees, ... | 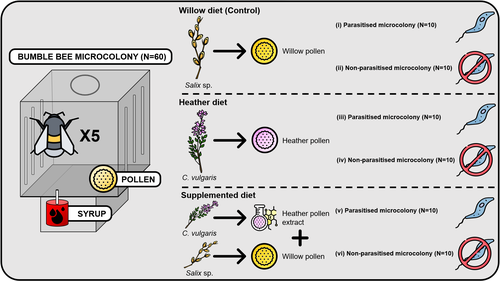 | Botany, Chemical ecology, Host-parasite interactions, Pollination, Zoology | Ignasi Bartomeus | | 2023-04-10 21:22:34 | View |
Interplay between historical and current features of the cityscape in shaping the genetic structure of the house mouse (Mus musculus domesticus) in Dakar (Senegal, West Africa)
Claire Stragier, Sylvain Piry, Anne Loiseau, Mamadou Kane, Aliou Sow, Youssoupha Niang, Mamoudou Diallo, Arame Ndiaye, Philippe Gauthier, Marion Borderon, Laurent Granjon, Carine Brouat, Karine Berthier
https://doi.org/10.1101/557066
Urban past predicts contemporary genetic structure in city rats
Recommended by Michelle DiLeo based on reviews by Torsti Schulz, ? and 1 anonymous reviewer
Urban areas are expanding worldwide, and have become a dominant part of the landscape for many species. Urbanization can fragment pre-existing populations of vulnerable species leading to population declines and the loss of connectivity. On the other hand, expansion of urban areas can also facilitate the spread of human commensals including pests. Knowledge of the features of cityscapes that facilitate gene flow and maintain diversity of pests is thus key to their management and eradication.
Cities are complex mosaics of natural and manmade surfaces, and habitat quality is not only influenced by physical aspects of the cityscape but also by socioeconomic factors and human behaviour. Constant development means that cities also change rapidly in time; contemporary urban life reflects only a snapshot of the environmental conditions faced by populations. It thus remains a challenge to identify the features that actually drive ecology and evolution of populations in cities [1]. While several studies have highlighted strong urban clines in genetic structure and adaption [2], few have considered the influence of factors beyond physical aspects of the cityscape or historical processes.
In this paper, Stragier et al. [3] sought to identify the current and past features of the cityscape and socioeconomic factors that shape genetic structure and diversity of the house mouse (Mus musculus domesticus) in Dakar, Senegal. The authors painstakingly digitized historical maps of Dakar from the time of European settlement in 1862 to present. The authors found that the main spatial genetic cline was best explained by historical cityscape features, with higher apparent gene flow and genetic diversity in areas that were connected earlier to initial European settlements. Beyond the main trend of spatial genetic structure, they found further evidence that current features of the cityscape were important. Specifically, areas with low vegetation and poor housing conditions were found to support large, genetically diverse populations. The authors demonstrate that their results are reproducible using several statistical approaches, including modeling that explicitly accounts for spatial autocorrelation.
The work of Stragier et al. [3] thus highlights that populations of city-dwelling species are the product of both past and present cityscapes. Going forward, urban evolutionary ecologists should consider that despite the potential for rapid evolution in urban landscapes, the signal of a species’ colonization can remain for generations.
References
[1] Rivkin, L. R., Santangelo, J. S., Alberti, M. et al. (2019). A roadmap for urban evolutionary ecology. Evolutionary Applications, 12(3), 384-398. doi: 10.1111/eva.12734
[2] Miles, L. S., Rivkin, L. R., Johnson, M. T., Munshi‐South, J. and Verrelli, B. C. (2019). Gene flow and genetic drift in urban environments. Molecular ecology, 28(18), 4138-4151. doi: 10.1111/mec.15221
[3] Stragier, C., Piry, S., Loiseau, A., Kane, M., Sow, A., Niang, Y., Diallo, M., Ndiaye, A., Gauthier, P., Borderon, M., Granjon, L., Brouat, C. and Berthier, K. (2020). Interplay between historical and current features of the cityscape in shaping the genetic structure of the house mouse (Mus musculus domesticus) in Dakar (Senegal, West Africa). bioRxiv, 557066, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/557066
| Interplay between historical and current features of the cityscape in shaping the genetic structure of the house mouse (Mus musculus domesticus) in Dakar (Senegal, West Africa) | Claire Stragier, Sylvain Piry, Anne Loiseau, Mamadou Kane, Aliou Sow, Youssoupha Niang, Mamoudou Diallo, Arame Ndiaye, Philippe Gauthier, Marion Borderon, Laurent Granjon, Carine Brouat, Karine Berthier | <p>Population genetic approaches may be used to investigate dispersal patterns of species living in highly urbanized environment in order to improve management strategies for biodiversity conservation or pest control. However, in such environment,... | 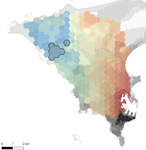 | Biological invasions, Landscape ecology, Molecular ecology | Michelle DiLeo | | 2019-02-22 08:36:13 | View |
Sexual segregation in a highly pagophilic and sexually dimorphic marine predator
Christophe Barbraud, Karine Delord, Akiko Kato, Paco Bustamante, Yves Cherel
https://doi.org/10.1101/472431
Sexual segregation in a sexually dimorphic seabird: a matter of spatial scale
Recommended by Denis Réale based on reviews by Dries Bonte and 1 anonymous reviewer
Sexual segregation appears in many taxa and can have important ecological, evolutionary and conservation implications. Sexual segregation can take two forms: either the two sexes specialise in different habitats but share the same area (habitat segregation), or they occupy the same habitat but form separate, unisex groups (social segregation) [1,2]. Segregation would have evolved as a way to avoid, or at least, reduce intersexual competition.
Testing whether social or habitat segregation is at play necessitates the use of combined approaches to determine the spatial scale at which segregation occurs. This enterprise is even more challenging when studying marine species, which travel over long distances to reach their foraging areas. This is what Barbraud et al. [3] have endeavoured on the snow petrel (Pagodroma nivea), a sexually dimorphic, polar seabird. Studying sexual segregation at sea requires tools for indirect measures of habitat use and foraging tactics. During the incubation period, in a colony based at Pointe Geologie, Adelie land, East Antarctica, the team has equipped birds with GPS loggers to analyse habitat use and foraging behaviour. It has also compared short-, mid-, and long-term stable isotopic profiles, from plasma, blood cells, and feather samples, respectively.
Barbraud et al. [3] could not detect any evidence for sexual segregation in space use. Furthermore, the two sexes showed similar δ13C profiles, illustrating similar foraging latitudes, and indicating no sexual segregation at large spatial scales. Snow petrels all forage exclusively in the sea ice environment formed over the deep Antarctic continental shelf. The authors, however, found other forms of segregation: males consistently foraged at higher sea ice concentrations than females. Males also fed on higher trophic levels than females. Therefore, male and female snow petrels segregate at a smaller spatial scale, and use different foraging tactics and diet specialisations. Females also took shorter foraging trips than males, with higher mass gain that strongly benefit from higher sea ice concentration. Mass gain in males increased with the length of their foraging trip at sea ice areas.
The authors conclude that high sea ice concentration offers the most favourable foraging habitat for snow petrels, and thus that intersexual competition may drive females away from high sea ice areas. This study shows that combining information from different tools provides an elegant way of isolating the potential factors driving sexual segregation and the spatial scales at which it occurs.
References
[1] Conradt, L. (2005). Definitions, hypotheses, models and measures in the study of animal segregation. In Sexual segregation in vertebrates: ecology of the two sexes (Ruckstuhl K.E. and Neuhaus, P. eds). Cambridge University Press, Cambridge, United Kingdom. Pp:11–34.
[2] Ruckstuhl, K. E. (2007). Sexual segregation in vertebrates: proximate and ultimate causes. Integrative and Comparative Biology, 47(2), 245-257. doi: 10.1093/icb/icm030
[3] Barbraud, C., Delord, K., Kato, A., Bustamante, P., & Cherel, Y. (2018). Sexual segregation in a highly pagophilic and sexually dimorphic marine predator. bioRxiv, 472431, ver. 3 peer-reviewed and recommended bt PCI Ecology. doi: 10.1101/472431
| Sexual segregation in a highly pagophilic and sexually dimorphic marine predator | Christophe Barbraud, Karine Delord, Akiko Kato, Paco Bustamante, Yves Cherel | <p>Sexual segregation is common in many species and has been attributed to intra-specific competition, sex-specific differences in foraging efficiency or in activity budgets and habitat choice. However, very few studies have simultaneously quantif... |  | Foraging, Marine ecology | Denis Réale | Dries Bonte, Anonymous | 2018-11-19 13:40:59 | View |
Trophic cascade driven by behavioural fine-tuning as naïve prey rapidly adjust to a novel predator
Chris J Jolly, Adam S Smart, John Moreen, Jonathan K Webb, Graeme R Gillespie and Ben L Phillips
https://doi.org/10.1101/856997
While the quoll’s away, the mice will play… and the seeds will pay
Recommended by Denis Réale based on reviews by 2 anonymous reviewers
A predator can strongly influence the demography of its prey, which can have profound carryover effects on the trophic network; so-called density-mediated indirect interactions (DMII; Werner and Peacor 2003; Schmitz et al. 2004; Trussell et al. 2006). Furthermore, a novel predator can alter the phenotypes of its prey for traits that will change prey foraging efficiency. These trait-mediated indirect interactions may in turn have cascading effects on the demography and features of the basal resources consumed by the intermediate consumer (TMIII; Werner and Peacor 2003; Schmitz et al. 2004; Trussell et al. 2006), but very few studies have looked for these effects (Trusell et al. 2006). The study “Trophic cascade driven by behavioural fine-tuning as naïve prey rapidly adjust to a novel predator”, by Jolly et al. (2020) is therefore a much-needed addition to knowledge in this field. The authors have profited from a rare introduction of Northern quolls (Dasyurus hallucatus) on an Australian island, to examine both the density-mediated and trait-mediated indirect interactions with grassland melomys (Melomys burtoni) and the vegetation of their woodland habitat.
Jolly et al. (2020) compared melomys populations in four quoll-invaded and three quoll-free sites on the same island. Using capture-mark-recapture methods, they found a lower survival and decreased population size in quoll-invaded sites compared to quoll-free sites. Although they acknowledge that this decline could be attributable to either the direct effects of the predator or to a wildfire that occurred early in the experiment in the quoll-invaded sites, the authors argue that the wildfire alone cannot explain all of their results.
Beyond demographic effects, Jolly et al. (2020) also examined risk taking, foraging behaviour, and predator avoidance in melomys. Quoll presence was first associated with a strong decrease in risk taking in melomys, but the difference disappeared over the three years of study, indicating a possible adjustment by the prey. In quoll-invaded sites, though, melomys continued to be more neophobic than in the quoll-free sites throughout the study. Furthermore, in a seed (i.e. wheat) removal experiment, Jolly et al. (2020) measured how melomys harvested seeds in the presence or absence of predator scents. In both quoll-invaded and quoll-free sites, melomys density increased seed harvest efficiency. Melomys also removed less seeds in quoll-invaded sites than in quoll-free sites, supporting both the DMII and TMII hypotheses. However, in the quoll-invaded sites only, melomys foraged less on predator-scented seed patches than on unscented ones, trading foraging efficiency for an increased safety against predators, and this effect increased across the years. This last result indicates that predators can indirectly influence seed consumption through the trade-off between foraging and predator avoidance, strongly supporting the TMII hypothesis.
Ideally, the authors would have run a nice before-after, impact-control design, but nature does not always allow for ideal experimental designs. Regardless, the results of such an “experiment in the wild” predation study are still valuable, as they are very rare (Trussell et al. 2006), and they provide crucial information on the direct and indirect interactions along a trophic cascade. Furthermore, the authors have effectively addressed any concerns about potential confounding factors, and thus have a convincing argument that their results represent predator-driven demographic and behavioural changes.
One important question remains from an evolutionary ecology standpoint: do the responses of melomys to the presence of quolls represent phenotypically plastic changes or rapid evolutionary changes caused by novel selection pressures? Classically, TMII are assumed to be mostly caused by phenotypic plasticity (Werner and Peacor 2003), and this might be the case when the presence of the predator is historical. Phenotypic plasticity allows quick and reversible adjustments of the prey population to changes in the predator density. When the predator population declines, such rapid phenotypic changes can be reversed, reducing the cost associated with anti-predator behaviour (e.g., lower foraging efficiency) in the absence of predators. In the case of a novel predator, however, short-term evolutionary responses by the prey may play role in the TMII, as they would allow a phenotypic shift in prey’s traits along the trade-off between foraging efficiency and anti-predator response that will probably more advantageous over the longer term, if the predator does not disappear. The authors state that they could not rule out one or the other of these hypotheses. However, future work estimating the relative importance of phenotypic plasticity and evolutionary changes in the quoll-melomys system would be valuable. Phenotypic selection analysis, for example, by estimating the link between survival and the traits measured, might help test for a fitness advantage to altered behaviour in the presence of a predator. Common garden experiments, comparing the quoll-invaded and the quoll-free melomys populations, might also provide information on any potential evolutionary changes caused by predation. More work could also analyse the potential effects on the seed populations. Not only might the reduction in seed predation have consequences on the landscape in the future, as the authors mention, but it may also mean that the seeds themselves could be subject to novel selection pressures, which may affect their phenology, physiology or life history. Off course, the authors will have to switch from wheat to a more natural situation, and evaluate the effects of changes in the melomys population on the feature of the local vegetation and the ecosystem.
Finally, the authors have not yet found that the observed changes in the traits have translated into a demographic rebound for melomys. Here again, I can see an interesting potential for further studies. Should we really expect an evolutionary rescue (Bell and Gonzalez 2009) in this system? Alternatively, should the changes in behaviour be accompanied by permanent changes in life history, such as a slower pace-of-life (Réale et al. 2010) that could possibly lead to lower melomys density?
This paper provides nice in natura evidence for density- and trait-mediated indirect interactions hypotheses. I hope it will be the first of a long series of work on this interesting quoll-melomys system, and that the authors will be able to provide more information on the eco-evolutionary consequences of a novel predator on a trophic network.
References
-Bell G, Gonzalez A (2009) Evolutionary rescue can prevent extinction following environmental change. Ecology letters, 12(9), 942-948. https://doi.org/10.1111/j.1461-0248.2009.01350.x
-Jolly CJ, Smart AS, Moreen J, Webb JK, Gillespie GR, Phillips BL (2020) Trophic cascade driven by behavioural fine-tuning as naïve prey rapidly adjust to a novel predator. bioRxiv, 856997, ver. 6 peer-reviewed and recommended by PCI Ecology. https://doi.org/ 10.1101/856997
-Matassa C, Ewanchuk P, Trussell G (2018) Cascading effects of a top predator on intraspecific competition at intermediate and basal trophic levels. Functional Ecology, 32(9), 2241-2252. https://doi.org/10.1111/1365-2435.13131
-Réale D, Garant D, Humphries MM, Bergeron P, Careau V, Montiglio PO (2010) Personality and the emergence of the pace-of-life syndrome concept at the population level. Philosophical Transactions of the Royal Society B: Biological Sciences, 365(1560), 4051-4063. https://doi.org/10.1098/rstb.2010.0208
-Schmitz O, Krivan V, Ovadia O (2004) Trophic cascades: the primacy of trait‐mediated indirect interactions. Ecology Letters 7(2), 153-163. https://doi.org/10.1111/j.1461-0248.2003.00560.x
-Trussell G, Ewanchuk P, Matassa C (2006). Habitat effects on the relative importance of trait‐ and density‐mediated indirect interactions. Ecology Letters, 9(11), 1245-1252. https://doi.org/10.1111/j.1461-0248.2006.00981.x
-Werner EE, Peacor SD (2003) A review of trait‐mediated indirect interactions in ecological communities. Ecology, 84(5), 1083-1100. https://doi.org/10.1890/0012-9658(2003)084[1083:AROTII]2.0.CO;2
| Trophic cascade driven by behavioural fine-tuning as naïve prey rapidly adjust to a novel predator | Chris J Jolly, Adam S Smart, John Moreen, Jonathan K Webb, Graeme R Gillespie and Ben L Phillips | <p>The arrival of novel predators can trigger trophic cascades driven by shifts in prey numbers. Predators also elicit behavioural change in prey populations, via phenotypic plasticity and/or rapid evolution, and such changes may also contribute t... |  | Behaviour & Ethology, Biological invasions, Evolutionary ecology, Experimental ecology, Foraging, Herbivory, Population ecology, Terrestrial ecology, Tropical ecology | Denis Réale | | 2019-11-27 21:39:44 | View |
Does elevated parasite richness in the environment affect daily path length of animals or is it the converse? An answer bringing some new elements of discussion
Recommended by Cédric Sueur based on reviews by 2 anonymous reviewers
In 2015, Brockmeyer et al. [1] suggested that mandrills (Mandrillus sphinx) may accept additional ranging costs to avoid heavily parasitized areas. Following this paper, Bicca-Marques and Calegaro-Marques [2] questioned this interpretation and presented other hypotheses. To summarize, whilst Brockmeyer et al. [1] proposed that elevated daily path length may be a consequence of elevated parasite richness, Bicca-Marques and Calegaro-Marques [2] viewed it as a cause. In this current paper, Charpentier and Kappeler [3] respond to some of the criticisms by Bicca-Marques and Calegaro-Marques and discuss the putative parsimony of the two competing scenarios. The manuscript is interesting and focuses on an important question concerning the discussion about the social organization and home range use in wild mandrills. This answer helps to move this debate forward and should stimulate more empirical studies of the role of environmentally-transmitted parasites in shaping ranging and movement patterns of wild vertebrates. Given the elements this paper brings to the topics, it should have been published in American Journal of Primatology, the journal that published the two previous articles.
References
[1] Brockmeyer, T., Kappeler, P. M., Willaume, E., Benoit, L., Mboumba, S., & Charpentier, M. J. E. (2015). Social organization and space use of a wild mandrill (Mandrillus sphinx) group. American Journal of Primatology, 77(10), 1036–1048. doi: 10.1002/ajp.22439
[2] Bicca-Marques, J. C., & Calegaro-Marques, C. (2016). Ranging behavior drives parasite richness: A more parsimonious hypothesis. American Journal of Primatology, 78(9), 923–927. doi: 10.1002/ajp.22561
[3] Charpentier, M. J., & Kappeler, P. M. (2018). A reply to “Ranging Behavior Drives Parasite Richness: A More Parsimonious Hypothesis.” ArXiv:1805.08151v2 [q-Bio]. Retrieved from http://arxiv.org/abs/1805.08151
| A reply to “Ranging Behavior Drives Parasite Richness: A More Parsimonious Hypothesis” | Charpentier MJE, Kappeler PM | In a recent article, Bicca-Marques and Calegaro-Marques [2016] discussed the putative assumptions related to an interpretation we provided regarding an observed positive relationship between weekly averaged parasite richness of a group of mandrill... |  | Behaviour & Ethology, Evolutionary ecology, Foraging, Host-parasite interactions, Spatial ecology, Metacommunities & Metapopulations, Zoology | Cédric Sueur | | 2018-05-22 10:59:33 | View |
Assessing bat-vehicle collision risks using acoustic 3D tracking
Recommended by Gloriana Chaverri based on reviews by Mark Brigham and ?
The loss of biodiversity is an issue of great concern, especially if the extinction of species or the loss of a large number of individuals within populations results in a loss of critical ecosystem services. We know that the most important threat to most species is habitat loss and degradation (Keil et al., 2015; Pimm et al., 2014); the latter can be caused by multiple anthropogenic activities, including pollution, introduction of invasive species and fragmentation (Brook et al., 2008; Scanes, 2018). Roads are a major cause of habitat fragmentation, isolating previously connected populations and being a direct source of mortality for animals that attempt to cross them (Spellberg, 1998).
While most studies have focused on the effect of roads on larger mammals (Bartonička et al., 2018; Litvaitis and Tash, 2008), in recent years many researchers have grown increasingly concerned about the risk of collision between bats and vehicles (Fensome and Mathews, 2016). For example, a recent publication by Medinas et al. (2021) found 509 bat casualties along a 51-km-long transect during a period of 3 years. Their study provides extremely valuable information to asses which factors primarily drive bat mortality on roads, yet it required a substantial investment of time coupled with the difficulty of detecting bat carcasses. Other studies have used acoustic monitoring as a proxy to gauge risk of collision based on estimates of bat density along roads (reviewed in Fensome and Mathews 2016); while the results of such studies are valuable, the number of passes recorded does not necessarily equal collision risk, as many species may simply avoid crossing the roads. Understanding the risk of collisions is of vital importance for adequate planning of road construction, particularly for key sites that harbor threatened bat species or unusually large populations, especially if these are already greatly impacted by other anthropogenic activities (e.g. wind turbines; Kunz et al. 2007) or unusually deadly pathogens (e.g. white-nose syndrome; Blehert et al. 2009).
The study by Roemer et al. (2020) titled “Influence of local landscape and time of year on bat-road collision risks”, is a welcome addition to our understanding of bat collision risk as it employs a more accurate assessment of bat collision risk based on acoustic monitoring and tracking of flight paths. The goal of the study of Roemer and collaborators, which was conducted at 66 study sites in the Mediterranean region, is to provide an assessment of collision risk based on bat activity near roads. They collected a substantial amount of information for several species: more than 30,000 estimated flight trajectories for 21+ species, including Barbastella barbastellus, Myotis spp., Plecotus sp., Rhinolophus ferrumequinum, Miniopterus schreibersii, Pipistrellus spp., Nyctalus leisleri, and others. They assess risk based on estimates of 1) species abundance from acoustic monitoring, 2) direction of flight paths along roads, and 3) bat-vehicle co-occurrence.
Their findings suggest that risk is habitat, species, guild, and season-specific. Roads within forested habitats posed the largest threats for most species, particularly since most flights within these habitats occurred at the zone of collision risk. They also found that bats typically fly parallel to the road axis regardless of habitat type, which they argue supports the idea that bats may use roads as corridors. The results of their study, as expected, also show that the majority of bat passes were detected during summer or autumn, depending on species, yet they provide novel findings of an increase in risky behaviors during autumn, when the number of passes at the zone of collision risk increased significantly. Their results also suggest that mid-range echolocators, a classification that is based on call design and parameters (Frey-Ehrenbold et al., 2013), had a larger portion of flights in the zone at risk, thus potentially making them more susceptible than short and long-range echolocators to collisions with vehicles.
The methods employed by Roemer et al. (2020) could further help us determine how roads pose species and site-specific threats in a diversity of places without the need to invest a significant amount of time locating bat carcasses. Their findings are also important as they could provide valuable information for deciding where new roads should be constructed, particularly if the most vulnerable species are abundant, perhaps due to the presence of important roost sites. They also show how habitats near larger roads could increase threats, providing an important first step for recommendations regarding road construction and maintenance. As pointed out by one reviewer, one possible limitation of the study is that the results are not supported by the identification of carcasses. For example, does an increase in the number of identified flights at the zone of risk really translate into an increase in the number of collisions? Regardless of the latter, the paper’s methods and results are very valuable and provide an important step towards developing additional tools to assess bat-vehicle collision risks.
References
[1] Bartonička T, Andrášik R, Duľa M, Sedoník J, Bíl M (2018) Identification of local factors causing clustering of animal-vehicle collisions. The Journal of Wildlife Management, 82, 940–947. https://doi.org/10.1002/jwmg.21467
[2] Blehert DS, Hicks AC, Behr M, Meteyer CU, Berlowski-Zier BM, Buckles EL, Coleman JTH, Darling SR, Gargas A, Niver R, Okoniewski JC, Rudd RJ, Stone WB (2009) Bat White-Nose Syndrome: An Emerging Fungal Pathogen? Science, 323, 227–227. https://doi.org/10.1126/science.1163874
[3] Brook BW, Sodhi NS, Bradshaw CJA (2008) Synergies among extinction drivers under global change. Trends in Ecology & Evolution, 23, 453–460. https://doi.org/10.1016/j.tree.2008.03.011
[4] Fensome AG, Mathews F (2016) Roads and bats: a meta-analysis and review of the evidence on vehicle collisions and barrier effects. Mammal Review, 46, 311–323. https://doi.org/10.1111/mam.12072
[5] Frey‐Ehrenbold A, Bontadina F, Arlettaz R, Obrist MK (2013) Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matrices. Journal of Applied Ecology, 50, 252–261. https://doi.org/10.1111/1365-2664.12034
[6] Keil P, Storch D, Jetz W (2015) On the decline of biodiversity due to area loss. Nature Communications, 6, 8837. https://doi.org/10.1038/ncomms9837
[7] Kunz TH, Arnett EB, Erickson WP, Hoar AR, Johnson GD, Larkin RP, Strickland MD, Thresher RW, Tuttle MD (2007) Ecological impacts of wind energy development on bats: questions, research needs, and hypotheses. Frontiers in Ecology and the Environment, 5, 315–324. https://doi.org/10.1890/1540-9295(2007)5[315:EIOWED]2.0.CO;2
[8] Litvaitis JA, Tash JP (2008) An Approach Toward Understanding Wildlife-Vehicle Collisions. Environmental Management, 42, 688–697. https://doi.org/10.1007/s00267-008-9108-4
[9] Medinas D, Marques JT, Costa P, Santos S, Rebelo H, Barbosa AM, Mira A (2021) Spatiotemporal persistence of bat roadkill hotspots in response to dynamics of habitat suitability and activity patterns. Journal of Environmental Management, 277, 111412. https://doi.org/10.1016/j.jenvman.2020.111412
[10] Pimm SL, Jenkins CN, Abell R, Brooks TM, Gittleman JL, Joppa LN, Raven PH, Roberts CM, Sexton JO (2014) The biodiversity of species and their rates of extinction, distribution, and protection. Science, 344. https://doi.org/10.1126/science.1246752
[11] Roemer C, Coulon A, Disca T, Bas Y (2020) Influence of local landscape and time of year on bat-road collision risks. bioRxiv, 2020.07.15.204115, ver. 3 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2020.07.15.204115
[12] Scanes CG (2018) Chapter 19 - Human Activity and Habitat Loss: Destruction, Fragmentation, and Degradation. In: Animals and Human Society (eds Scanes CG, Toukhsati SR), pp. 451–482. Academic Press. https://doi.org/10.1016/B978-0-12-805247-1.00026-5
[13] Spellerberg I (1998) Ecological effects of roads and traffic: a literature review. Global Ecology & Biogeography Letters, 7, 317–333. https://doi.org/10.1046/j.1466-822x.1998.00308.x
| Influence of local landscape and time of year on bat-road collision risks | Charlotte Roemer, Aurélie Coulon, Thierry Disca, and Yves Bas | <p>Roads impact bat populations through habitat loss and collisions. High quality habitats particularly increase bat mortalities on roads, yet many questions remain concerning how local landscape features may influence bat behaviour and lead to hi... |  | Behaviour & Ethology, Biodiversity, Conservation biology, Human impact, Landscape ecology | Gloriana Chaverri | | 2020-07-20 10:56:29 | View |
Use of linear features by red-legged partridges in an intensive agricultural landscape: implications for landscape management in farmland
Charlotte Perrot, Antoine Berceaux, Mathias Noel, Beatriz Arroyo, Leo Bacon
https://doi.org/10.1101/2023.07.27.550774
The importance of managing linear features in agricultural landscapes for farmland birds
Recommended by Ricardo Correia based on reviews by Matthew Grainger and 1 anonymous reviewer
European farmland bird populations continue declining at an alarming rate, and some species require urgent action to avoid their demise (Silva et al. 2024). While factors such as climate change and urbanization also play an important role in driving the decline of farmland bird populations, its main driver seems to be linked with agricultural intensification (Rigal et al. 2023). Besides increased pesticide and fertilizer use, agricultural intensification often results in the homogenization of agricultural landscapes through the removal of seminatural linear features such as hedgerows, field margins, and grassy strips that can be beneficial for biodiversity. These features may be particularly important during the breeding season, when breeding farmland birds can benefit from patches of denser vegetation to conceal nests and improve breeding success. It is both important and timely to understand how landscape management can help to address the ongoing decline of farmland birds by identifying specific actions that can boost breeding success.
Perrot et al. 2023 contribute to this effort by exploring how red-legged partridges use linear features in an intensive agricultural landscape during the breeding season. Through a combination of targeted fieldwork and GPS tracking, the authors highlight patterns in home range size and habitat selection that provide insights for landscape management. Specifically, their results suggest that birds have smaller range sizes in the vicinity of traffic routes and seminatural features structured by both herbaceous and woody cover. Furthermore, they show that breeding birds tend to choose linear elements with herbaceous cover whereas non-breeders prefer linear elements with woody cover, underlining the importance of accounting for the needs of both breeding and non-breeding birds. In particular, the authors stress the importance of providing additional vegetation elements such as hedges, grassy strips or embankments in order to increase landscape heterogeneity. These landscape elements are usually found in the vicinity of linear infrastructures such as roads and tracks, but it is important they are available also in separate areas to avoid the risk of bird collision and the authors provide specific recommendations towards this end. Overall, this is an important study with clear recommendations on how to improve landscape management for these farmland birds.
References
Perrot, C., Séranne, L., Berceaux, A., Noel, M., Arroyo, B., & Bacon, L. (2023) "Use of linear features by red-legged partridges in an intensive agricultural landscape: implications for landscape management in farmland." bioRxiv, ver. 2 peer-reviewed and recommended by Peer Community in Ecology.
https://doi.org/10.1101/2023.07.27.550774
Rigal, S., Dakos, V., Alonso, H., Auniņš, A., Benkő, Z., Brotons, L., ... & Devictor, V. (2023) "Farmland practices are driving bird population decline across Europe." Proceedings of the National Academy of Sciences 120.21: e2216573120.
https://doi.org/10.1073/pnas.2216573120
Silva, J. P., Gameiro, J., Valerio, F., & Marques, A. T. (2024) "Portugal's farmland bird crisis requires action." Science 383.6679: 157-157.
https://doi.org/10.1126/science.adn1390
| Use of linear features by red-legged partridges in an intensive agricultural landscape: implications for landscape management in farmland | Charlotte Perrot, Antoine Berceaux, Mathias Noel, Beatriz Arroyo, Leo Bacon | <p>Current agricultural practices and change are the major cause of biodiversity loss. An important change associated with the intensification of agriculture in the last 50 years is the spatial homogenization of the landscape with substantial loss... |  | Agroecology, Behaviour & Ethology, Biodiversity, Conservation biology, Habitat selection | Ricardo Correia | | 2023-08-01 10:27:33 | View |







