Diapause is not selected as a bet-hedging strategy in insects: a meta-analysis of reaction norm shapes
Jens Joschinski and Dries Bonte
10.1101/752881
When to diapause or not to diapause? Winter predictability is not the answer
Recommended by Bastien Castagneyrol based on reviews by Kévin Tougeron, Md Habibur Rahman Salman and 1 anonymous reviewer
Winter is a harsh season for many organisms that have to cope with food shortage and potentially lethal temperatures. Many species have evolved avoidance strategies. Among them, diapause is a resistance stage many insects use to overwinter. For an insect, it is critical to avoid lethal winter temperatures and thus to initiate diapause before winter comes, while making the most of autumn suitable climatic conditions [1,2]. Several cues can be used to appreciate that winter is coming, including day length and temperature [3]. But climate changes, temperatures rise and become more variable from year to year, which imposes strong pressure upon insect phenology [4]. How can insects adapt to changes in the mean and variance of winter onset?
In this paper, Jens Joschinski and Dries Bonte [5] address this question by using a well conducted meta-analysis of 458 diapause reaction norms obtained from 60 primary studies. They first ask first if insect mean diapause timing is tuned to match winter onset. They further ask if insects adapt to climatic unpredictability through a bet-hedging strategy by playing it safe and avoid risk (conservative bet-hedging) or on the contrary by avoiding to put all their eggs in one basket and spread the risk among their offspring (diversified bet-hedging). From published papers, the authors extracted data on mean diapause timing and information on latitude from which they retrieved day length inducing diapause, the date of winter onset and the day length at winter onset.
They found a positive correlation between latitude and the day length inducing diapause. On the contrary they found positive but (very) weak correlation between the date of winter onset and the date of diapause, thus indicating that diapause timing is not as optimally adapted to local environments as expected, particularly at high latitudes. They only found weak correlations between climate unpredictability and variability in diapause timing, and no correlation between climate unpredictability and deviation from optimal diapause timing. Together, these findings go against the hypothesis that insects use diversified or conservative bet-hedging strategies to cope with uncertainty in climatic conditions.
This is what makes the study thought provoking: the results do not match the theory well. Not because of a lack of data or a narrow scope, but because diapause is a complex trait that is determined by a large array of physiological and ecological factors [3]. Determining what are these factors is of particular interest in the face of the current climate change. This study shows what does not determine the timing of insect diapause. Researchers now know where to look at to improve our understanding of this key aspect of insect adaptation to climatic conditions.
References
[1] Dyck, H. V., Bonte, D., Puls, R., Gotthard, K., and Maes, D. (2015). The lost generation hypothesis: could climate change drive ectotherms into a developmental trap? Oikos, 124(1), 54–61. doi: 10.1111/oik.02066
[2] Gallinat, A. S., Primack, R. B., and Wagner, D. L. (2015). Autumn, the neglected season in climate change research. Trends in Ecology & Evolution, 30(3), 169–176. doi: 10.1016/j.tree.2015.01.004
[3] Tougeron, K. (2019). Diapause research in insects: historical review and recent work perspectives. Entomologia Experimentalis et Applicata, 167(1), 27–36. doi: 10.1111/eea.12753
[4] Bale, J. S., and Hayward, S. a. L. (2010). Insect overwintering in a changing climate. Journal of Experimental Biology, 213(6), 980–994. doi: 10.1242/jeb.037911
[5] Joschinski, J., and Bonte, D. (2020). Diapause is not selected as a bet-hedging strategy in insects: a meta-analysis of reaction norm shapes. BioRxiv, 752881, ver. 3 recommended and peer-reviewed by PCI Ecology. doi: 10.1101/752881
| Diapause is not selected as a bet-hedging strategy in insects: a meta-analysis of reaction norm shapes | Jens Joschinski and Dries Bonte | Many organisms escape from lethal climatological conditions by entering a resistant resting stage called diapause, and it is essential that this strategy remains optimally timed with seasonal change. Climate change therefore exerts selection press... | 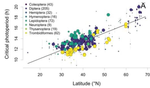 | Maternal effects, Meta-analyses, Phenotypic plasticity, Terrestrial ecology | Bastien Castagneyrol | | 2019-09-20 11:47:47 | View |
On the efficacy of restoration in stream networks: comments, critiques, and prospective recommendations
David Murray-Stoker
https://doi.org/10.1101/611939
A stronger statistical test of stream restoration experiments
Recommended by Karl Cottenie based on reviews by Eric Harvey and Mariana Perez Rocha
The metacommunity framework acknowledges that local sites are connected to other sites through dispersal, and that these connectivity patterns can influence local dynamics [1]. This framework is slowly moving from a framework that guides fundamental research to being actively applied in for instance a conservation context (e.g. [2]). Swan and Brown [3,4] analyzed the results of a suite of experimental manipulations in headwater and mainstem streams on invertebrate community structure in the context of the metacommunity concept. This was an important contribution to conservation ecology.
However, David Murray-Stoker [5] was not satisfied with their statistical analyses, and recreated, and more importantly, improved their original analyses in the peer-reviewed article. The new analyses are based on a combination of a more consistent site selection, checking the model assumptions, using different estimation procedures, and focusing more on effect size calculations versus statistical significance. This peer-reviewed article is thus the perfect example of the advantages of open research: the original authors making available both the data and their R script files, initially first updating the analyses and results themselves, followed by more in-depth analyses of the original data and question.
This peer reviewed went through a very in-depth process itself, with several rounds of questions and feedback that addressed both the statistical analyses, the interpretation of the results, and the conclusions. It also, however, addressed something that is often harder to provide feedback on, for instance the tone of the argument. I hope that scientists interested in these issues will not only read the final manuscript, but also the different steps of the peer review processes. These are very informative, I think, and provide a more complete picture of mainly the raison for certain decisions.
Not only does this provide the reader interested in stream conservation with the opportunity to make up their own mind on the appropriateness of these decisions, but it could potentially lead to more analyses of this important data set. For instance, maybe a formal meta-analysis that starts with the effect sizes of all the original studies might bring some new insights into this question?
References
[1] Leibold, M. A., Holyoak, M., Mouquet, N. et al. (2004). The metacommunity concept: a framework for multi‐scale community ecology. Ecology letters, 7(7), 601-613. doi: 10.1111/j.1461-0248.2004.00608.x
[2] Heino, J. (2013). The importance of metacommunity ecology for environmental assessment research in the freshwater realm. Biological Reviews, 88(1), 166-178. doi: 10.1111/j.1469-185X.2012.00244.x
[3] Swan, C. M., and Brown, B. L. (2017). Metacommunity theory meets restoration: isolation may mediate how ecological communities respond to stream restoration. Ecological Applications, 27(7), 2209-2219. doi: 10.1002/eap.1602
[4] Swan, C. M., and Brown, B. L. (2018). Erratum for: Metacommunity theory meets restoration: isolation may mediate how ecological communities respond to stream restoration. Ecological Applications 28:1370–1371. doi: 10.1002/eap.1738
[5] Murray-Stoker, D. (2020). On the efficacy of restoration in stream networks: comments, critiques, and prospective recommendations. bioRxiv, 611939, ver. 7 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/611939
| On the efficacy of restoration in stream networks: comments, critiques, and prospective recommendations | David Murray-Stoker | <p>Swan and Brown (2017) recently addressed the effects of restoration on stream communities under the meta-community framework. Using a combination of headwater and mainstem streams, Swan and Brown (2017) evaluated how position within a stream ne... |  | Community ecology, Freshwater ecology, Spatial ecology, Metacommunities & Metapopulations | Karl Cottenie | | 2019-09-21 22:12:57 | View |
Explore and move: a key to success in a changing world?
Recommended by Blandine Doligez based on reviews by Joe Nocera, Marion Nicolaus and Laure Cauchard
Changes in the spatial range of many species are one of the major consequences of the profound alteration of environmental conditions due to human activities. Some species expand, sometimes spectacularly during invasions; others decline; some shift. Because these changes result in local biodiversity loss (whether local species go extinct or are replaced by colonizing ones), understanding the factors driving spatial range dynamics appears crucial to predict biodiversity dynamics. Identifying the factors that shape individual movement is a main step towards such understanding. The study described in this preregistration (McCune et al. 2020) falls within this context by testing possible links between individual exploration behaviour and movements related to daily space use in an avian study model currently rapidly expanding, the great-tailed grackle (Quiscalus mexicanus).
Movement and exploration: which direction(s) for the link between exploration and dispersal?
Individuals are known to differ in their tendency to explore the environment (Réale et al. 2007; Wolf and Weissing 2012) and therefore in their motivation to move. Accordingly, exploration has been shown to relate to dispersal behaviour, i.e. movements between breeding sites (Dingemanse et al. 2003, Le Galliard et al. 2011, Rasmussen and Belk 2012; reviews in Cote et al. 2010, Ronce et al. 2012). Yet, the mechanisms underlying this link often remain unclear, due to the correlative nature of the data. A classical assumption is that dispersers may benefit from a high capacity to explore, allowing them to familiarize quicker with their new environment once reached, thus alleviating dispersal costs (Bonte et al. 2012). The association between dispersal and exploration would in this case result from selection for this combination of traits (Ronce et al. 2012), even though dispersal event itself may be independent from (and precede the effect of) exploration behaviour. Alternatively (but not exclusively), dispersal may simply be the final outcome of longer movements by individuals exploring larger ranges (Badyaev et al. 1996, Schliehe-Diecks et al. 2012). In the absence of easy ways to manipulate dispersal behaviour, on the one hand, and exploration tendency, on the other hand, investigating detailed, small-scale individual movements in relation to exploration should thus shed light on which processes may yield the observed relations between exploration as an individual personality trait and large-scale, long-term movements, such as dispersal, underlying species range dynamics.
In this project, the exploration behaviour of grackles will be measured in controlled conditions using standardized tests in captivity (McCune et al. 2019) before individuals are released and their daily space use behaviour will then be measured using remote tracking over long time periods (McCune et al. 2020). Importantly, these coupled measures will be obtained for individuals captured in three different populations: within the historical range of the species, in the middle of its expanding range and at the edge of the range (McCune et al. 2020). Therefore, the project will test (i) whether daily space use of individuals is linked to their intrinsic exploration tendency and (ii) whether space use differs between individuals from different populations along the expanding range. The preregistration echoes a complementary project by the same team that will focus on exploration and test (iii) whether exploration tendency differs between individuals from these different populations. Taken together, these three analyses will therefore provide solid background information to assess the role of exploration in the individuals’ decisions leading to movement and range dynamics in this species.
As underlined in the preregistration, previous studies addressing the links between individual exploration behaviour and movements have mostly focused on dispersal. A first type of studies have (as will be done here) measured exploration behaviour of individuals, often in captivity (Dingemanse et al. 2003, Korsten et al. 2013) but also in the wild (Rasmussen and Belk 2012, Debeffe et al. 2013), and related these measures to subsequent dispersal behaviour. The (often implicit) underlying assumption is that more exploratory individuals will be more likely to move further, explore different habitats and thus end up breeding farther than less explorative ones. In other words, exploration tendency precedes and drives dispersal. Sometimes, exploratory behaviour is measured on individuals of known dispersal status, i.e. after the dispersal event (Hoset et al. 2011), in which case selection for certain exploration phenotypes among dispersers may already have occurred. Besides this first approach, another type of studies have measured ‘exploration’ behaviour under the form of prospecting movements of individuals and linked these movements to subsequent dispersal (often in the context of habitat selection). While these studies were in the past based on direct thus potentially biased observations (Reed et al. 1999), they now rely more and more on technological advances using (miniaturized) remote tracking devices (Ponchon et al. 2013) that provide far more complete and unbiased movement data, and sometimes also complementary measures of individuals’ internal state. In this case, the implicit assumption is that individuals prospecting farther and/or in more habitat patches will be more likely to settle in a site located farther away from their departure site, because of a more exhaustive sampling of possible sites allowing individuals to identify higher-quality sites (Badyaev et al. 1996). In other words, exploration tendency would not directly lead to higher movements or longer distances, but would allow individuals to optimize their habitat choice among more numerous options, thus leading to an increased dispersal probability or distance; the relation between exploration and dispersal would thus be indirect. Prospecting studies address more closely the underlying mechanisms of movement; however, they cannot easily separate intrinsic individual exploratory tendency from the prospecting movements themselves, with potential feedback effects of the information already gathered on future exploration of other sites or patches, thus on subsequent movements.
By focusing on individual daily space use movements as a mechanistic approach to understand large-scale movements potentially involved in colonization and range expansion, the grackle study described in this preregistration (McCune et al. 2020) will thus contribute to bridge the knowledge gaps between exploration and dispersal. By linking exploration measures obtained from a battery of standardized tests conducted in controlled conditions to individual daily space use and movements recorded in the wild, the grackle project is set in between previous studies addressing the links between exploration and dispersal: it will document exploration in a separate and independent context with respect to the movements themselves, and it will use a mechanistic view of detailed movements by the same individuals in the wild to explore potential implications for dispersal and range expansion. Testing differences between the three study populations over the species range will indeed inform about potential large-scale, population implications of among-individual variation in the link between exploration and movements. Because this study will only measure already settled adult individuals whose previous history is unknown, there will nevertheless be no direct possible exploration of the link with either previous or subsequent dispersal behaviour. Thus, the potential links studied here relate more directly to post-dispersal benefits of exploration for an optimal exploitation of the new environment. Yet, if exploration is a life-long personality trait linked to daily movement patterns, it may also relate to natal dispersal movements in young individuals.
Evolutionary and conservation perspectives
If the results of the project reveal that exploration tendency and daily space use movements are indeed linked, and that individuals from populations across the species range differ in these traits, new questions will emerge. A first question would be whether such among-individual differences are at the origin of range expansion or rather one of its consequences since, again, we deal with correlative data here. In other words, individuals may differ in exploration tendency, and this may confer them different ability to move around, find and colonize new habitats; or individuals may show differences in exploration following arrival in a new habitat, either because more explorative individuals gain fitness benefits and are thus selected, or because of behavioural plasticity and post-colonization adjustment of exploration behaviour when facing new ecological and social conditions in the new environment. Another open question relates to the link between daily space use and dispersal: is dispersal a by-product of higher daily movements that allow individuals to discover new favorable places where to settle? Exploring this link could involve measuring just fledged individuals before natal dispersal occurs and/or individuals chosen according to their own dispersal history, and this would then imply long-term population monitoring as an efficient (but constraining) tool to address such questions. Finally, assessing the fitness consequences of the link between exploration and space use behaviour, and whether these consequences differ between populations along the range expansion, would also be needed to understand the contribution of this link to the invasion success of this species.
The study model chosen for this project is a rapidly expanding species. Importantly, however, and as emphasized in the preregistration, documenting links between exploration and daily space use patterns as well as differences between populations with different trajectories can provide crucial information in general to understand population persistence in response to global climate and landscape changes, both regarding invasion ability or extinction risk. The information should be key to assess the probability that a species may decline, persist or expand in studies addressing biodiversity and community dynamics in a changing world.
References
Badayev, A. V., Martin, T. E and Etges, W. J. 1996. Habitat sampling and habitat selection by female wild turkeys: ecological correlates and reproductive consequences. Auk 113: 636-646. doi: https://doi.org/10.2307/4088984
Bonte, D. et al. 2012. Costs of dispersal. Biological Reviews 87: 290-312. doi: https://doi.org/10.1111/j.1469-185X.2011.00201.x
Cote, J., Clobert, J., Brodin, T., Fogarty, S. and Sih, A. 2010. Personality-dependent dispersal: characterization, ontogeny and consequences for spatially structured populations. Philosophical Transactions of the Royal Society B 365: 4065-4576. doi: https://doi.org/10.1098/rstb.2010.0176
Debeffe, L., Morellet, N., Cargnelutti, B., Lourtet, B., Coulon, A., Gaillard, J.-M., Bon, R. and Hewison A. J. M. 2013. Exploration as a key component of natal dispersal: dispersers explore more than philopatric individuals in roe deer. Animal Behaviour 86: 143-151. doi: https://doi.org/10.1016/j.anbehav.2013.05.005
Dingemanse, N. J., Both, C., van Noordwijk, A. J., Rutten, A. L. and Drent, P. J. 2003. Natal dispersal and personalities in great tits (Parus major). Proceedings of the Royal Society B 270: 741-747. doi: https://doi.org/10.1098/rspb.2002.2300
Hoset, K. S., Ferchaud, A.-L., Dufour, F., Mersch, D., Cote, J. and Le Galliard, J.-F. 2011. Natal dispersal correlates with behavioral traits that are not consistent across early life stages. Behavioral Ecology 22: 176–183. doi: https://doi.org/10.1093/beheco/arq188
Korsten, P., van Overveld, T., Adriaensen, F. and Matthysen, E. 2013. Genetic integration of local dispersal and exploratory behaviour in a wild bird. Nature Communications 4: 2362. doi: https://doi.org/10.1038/ncomms3362
Le Galliard, J.-F., Rémy, A., Ims, R. A. and Lambin, X. 2011. Patterns and processes of dispersal behaviour in arvicoline rodents. Molecular Ecology 21: 505-523. doi: https://doi.org/10.1111/j.1365-294X.2011.05410.x
McCune K, Ross C, Folsom M, Bergeron L, Logan CJ. 2020. Does space use behavior relate to exploration in a species that is rapidly expanding its geographic range? http://corinalogan.com/Preregistrations/gspaceuse.html In principle acceptance by PCI Ecology of the version on 23 Sep 2020 https://github.com/corinalogan/grackles/blob/master/Files/Preregistrations/gspaceuse.Rmd.
McCune K, MacPherson M, Rowney C, Bergeron L, Folsom M, Logan CJ. 2019. Is behavioral flexibility linked with exploration, but not boldness, persistence, or motor diversity? (http://corinalogan.com/Preregistrations/gexploration.html) In principle acceptance by PCI Ecology of the version on 27 Mar 2019 https://github.com/corinalogan/grackles/blob/master/Files/Preregistrations/gexploration.Rmd
Ponchon, A., Grémillet, D., Doligez, B., Chambert, T., Tveraa, T., González-Solís, J. and Boulinier, T. 2013. Tracking prospecting movements involved in breeding habitat selection: insights, pitfalls and perspectives. Methods in Ecology and Evolution 4: 143-150. doi: https://doi.org/10.1111/j.2041-210x.2012.00259.x
Rasmussen, J. E. and Belk, M. C. 2012. Dispersal behavior correlates with personality of a North American fish. Current Zoology 58: 260–270. doi: https://doi.org/10.1093/CZOOLO%2F58.2.260
Réale, D., Reader, S. M., Sol, D., McDougall, P. T. and Dingemanse, N. J. 2007. Integrating animal temperament within ecology and evolution. Biological Reviews 82: 291-318. doi: https://doi.org/10.1111/j.1469-185x.2007.00010.x
Reed, J. M., Boulinier, T., Danchin, E. and Oring, L. W. 1999. Informed dispersal: prospecting by birds for breeding sites. Current Ornithology 15: 189-259. doi: https://doi.org/10.1007/978-1-4757-4901-4_5
Ronce, O. and Clobert, J. 2012. Dispersal syndromes. pp. 119-138 In Dispersal Ecology and Evolution (eds. Clobert, J., Baguette, M., Benton, T. G. and Bullock, J. M.), pp. 119-138. Oxford University Press.
Schliehe-Diecks, S., Eberle, M. and Kappeler, P. M. 2012. Walk the line - dispersal movements of gray mouse lemurs (Microcebus murinus). Behavioral Ecology and Sociobiology 66: 1175-1185. doi: https://dx.doi.org/10.1007%2Fs00265-012-1371-y
Wolf, M. and Weissing, F. J. 2012. Animal personalities: consequences for ecology and evolution. Trends in Ecology and Evolution 27: 452-461. doi: https://doi.org/10.1016/j.tree.2012.05.001
| Does space use behavior relate to exploration in a species that is rapidly expanding its geographic range? | Kelsey B. McCune, Cody Ross, Melissa Folsom, Luisa Bergeron, Corina Logan | Great-tailed grackles (Quiscalus mexicanus) are rapidly expanding their geographic range (Wehtje 2003). Range expansion could be facilitated by consistent behavioural differences between individuals on the range edge and those in other parts of th... |  | Behaviour & Ethology, Biological invasions, Conservation biology, Habitat selection, Phenotypic plasticity, Preregistrations, Spatial ecology, Metacommunities & Metapopulations | Blandine Doligez | | 2019-09-30 19:27:40 | View |
The persistence in time of distributional patterns in marine megafauna impacts zonal conservation strategies
Charlotte Lambert, Ghislain Dorémus, Vincent Ridoux
https://doi.org/10.1101/790634
The importance of spatio-temporal dynamics on MPA's design
Recommended by Sergio Estay based on reviews by Ana S. L. Rodrigues and 1 anonymous reviewer
Marine protected areas (MPA) have arisen as the main approach for conservation of marine species. Fishes, marine mammals and birds can be conservation targets that justify the implementation of these areas. However, MPAs undergo many of the problems faced by their terrestrial equivalent. One of the major concerns is that these conservation areas are spatially constrained, by logistic reasons, and many times these constraints caused that key areas for the species (reproductive sites, refugees, migration) fall outside the limits, making conservation efforts even more difficult. Lambert et al. [1] evaluate at what point the Bay of Biscay MPA contains key ecological areas for several emblematic species. The evaluation incorporated a spatio-temporal dimension. To evaluate these ideas, authors evaluate two population descriptors: aggregation and persistence of several species of cetaceans and seabirds.
The authors determined that despite the MPA contains key areas for some species, for many others the key areas fall outside the MPA (aggregation sites) or observed aggregation sites are poorly persistent in time. They found that aggregation and persistence behave as two uncorrelated descriptors of the spatio-temporal distribution of populations. Variability of both characteristics was species-specific, but in all cases the message is clear: both features must be taken into account to evaluate the effectiveness of MPAs. Both conclusions pointed out to the difficulties that a strategy based on MPAs could face when the target are those species with low aggregation or those where key sites show low persistence in time.
Conceptually, the manuscript and its conclusions are very interesting, specially its recommendation of including temporal variability of species abundances and aggregation in the design of MPAs. However, despite the clear biological importance of persistence and aggregation of the conservation targets for the design of a MPA, its implementation will still be an extremely complex task. A first constraint is that important areas for one species could not be relevant for others, making the design of the MPA difficult because the more target species we include the larger the area needed for the MPA. As a consequence, the management of the MPA turns difficult and expensive as the area increases. These increased costs could be a key point for accepting/rejecting the implementation of these MPAs for governments. Also larger areas could imply highest level of conflict with local communities or stakeholders. In many the inclusion inside MPAs of areas with traditional social or economic use will be a major source of conflict with the people.
Despite these difficulties, the results of Lambert et al. [1] give us a key message for improving MPA’s design. The best strategy for including their conclusions in the effective implementation of these areas will be the next target in conservation research.
References
[1] Lambert, C., Dorémus, G. and V. Ridoux (2020) The persistence in time of distributional patterns in marine megafauna impacts zonal conservation strategies. bioRxiv, 790634, ver. 3 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/790634
| The persistence in time of distributional patterns in marine megafauna impacts zonal conservation strategies | Charlotte Lambert, Ghislain Dorémus, Vincent Ridoux | <p>The main type of zonal conservation approaches corresponds to Marine Protected Areas (MPAs), which are spatially defined and generally static entities aiming at the protection of some target populations by the implementation of a management pla... |  | Conservation biology, Habitat selection, Species distributions | Sergio Estay | | 2019-10-03 08:47:17 | View |
A meta-analysis to infer generic predator functional response
Recommended by Samir Simon Suweis based on reviews by Ludek Berec and gyorgy barabas
Species interactions are classically derived from the law of mass action: the probability that, for example, a predation event occurs is proportional to the product of the density of the prey and predator species. In order to describe how predator and prey species populations grow, is then necessary to introduce functional response, describing the intake rate of a consumer as a function of food (e.g. prey) density.
Linear functional responses shapes are typically introduced in the ecological modeling of population dynamics for both predator-prey and mutualistic systems [1,2]. Recently some works have proposed alternatives to the classic approach for mutualistic systems [3,4], both because cooperative interactions also model effect not directly related to mass action [3] and for analytical tractability [4,5].
In this work [6] the authors challenge the classic modeling of functional response also for predator-prey systems. In particular, they use a meta-analysis of several observational studies of predator-prey ecosystems to infer a generic predator functional response, fitting a phenomenological generalization of the mass-action law. Using advanced statistical analysis, they show that the functional response obtained from data is clearly different from the mass-action assumption. In fact, they found that it scales sub-linearly as the square root of the ratio between predator and prey biomass. They further argue that, from a macro-ecological point of view, using such a phenomenological relationship might be more valuable than relying on various mechanistic functional response formulations.
The manuscript thus provides an interesting different perspective on how to approach predator-prey modelling and for this reason, I have recommended the work for PCI Ecology.
References
[1] Volterra, V. (1928). Variations and Fluctuations of the Number of Individuals in Animal Species living together. ICES Journal of Marine Science, 3(1), 3–51. doi: 10.1093/icesjms/3.1.3
[2] Bastolla, U., Fortuna, M. A., Pascual-García, A., Ferrera, A., Luque, B., and Bascompte, J. (2009). The architecture of mutualistic networks minimizes competition and increases biodiversity. Nature, 458(7241), 1018–1020. doi: 10.1038/nature07950
[3] Tu, C., Suweis, S., Grilli, J., Formentin, M., and Maritan, A. (2019). Reconciling cooperation, biodiversity and stability in complex ecological communities. Scientific Reports, 9(1), 1–10. doi: 10.1038/s41598-019-41614-2
[4] García-Algarra, J., Galeano, J., Pastor, J. M., Iriondo, J. M., and Ramasco, J. J. (2014). Rethinking the logistic approach for population dynamics of mutualistic interactions. Journal of Theoretical Biology, 363, 332–343. doi: 10.1016/j.jtbi.2014.08.039
[5] Suweis, S., Simini, F., Banavar, J. R., and Maritan, A. (2013). Emergence of structural and dynamical properties of ecological mutualistic networks. Nature, 500(7463), 449–452. doi: 10.1038/nature12438
[6] Barbier, M., Wojcik, L., and Loreau, M. (2020). A macro-ecological approach to predators’ functional response. BioRxiv, 832220, ver. 4 recommended and peer-reviewed by Peer Community in Ecology. doi: 10.1101/832220
| A macro-ecological approach to predators' functional response | Matthieu Barbier, Laurie Wojcik, Michel Loreau | <p>Predation often deviates from the law of mass action: many micro- and meso-scale experiments have shown that consumption saturates with resource abundance, and decreases due to interference between consumers. But does this observation hold at m... | 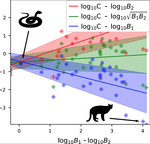 | Community ecology, Food webs, Meta-analyses, Theoretical ecology | Samir Simon Suweis | | 2019-11-08 15:42:16 | View |
Temperature predicts the maximum tree-species richness and water and frost shape the residual variation
Ricardo A. Segovia
https://doi.org/10.1101/836338
New light on the baseline importance of temperature for the origin of geographic species richness gradients
Recommended by Joaquín Hortal based on reviews by Rafael Molina-Venegas and 2 anonymous reviewers based on reviews by Rafael Molina-Venegas and 2 anonymous reviewers
Whether environmental conditions –in particular energy and water availability– are sufficient to account for species richness gradients (e.g. Currie 1991), or the effects of other biotic and historical or regional factors need to be considered as well (e.g. Ricklefs 1987), was the subject of debate during the 1990s and 2000s (e.g. Francis & Currie 2003; Hawkins et al. 2003, 2006; Currie et al. 2004; Ricklefs 2004). The metabolic theory of ecology (Brown et al. 2004) provided a solid and well-rooted theoretical support for the preponderance of energy as the main driver for richness variations. As any good piece of theory, it provided testable predictions about the sign and shape (i.e. slope) of the relationship between temperature –a key aspect of ambient energy– and species richness. However, these predictions were not supported by empirical evaluations (e.g. Kreft & Jetz 2007; Algar et al. 2007; Hawkins et al. 2007a), as the effects of a myriad of other environmental gradients, regional factors and evolutionary processes result in a wide variety of richness–temperature responses across different groups and regions (Hawkins et al. 2007b; Hortal et al. 2008). So, in a textbook example of how good theoretical work helps advancing science even if proves to be (partially) wrong, the evaluation of this aspect of the metabolic theory of ecology led to current understanding that, while species richness does respond to current climatic conditions, many other ecological, evolutionary and historical factors do modify such response across scales (see, e.g., Ricklefs 2008; Hawkins 2008; D’Amen et al. 2017). And the kinetic model linking mean annual temperature and species richness (Allen et al. 2002; Brown et al. 2004) was put aside as being, perhaps, another piece of the puzzle of the origin of current diversity gradients.
Segovia (2021) puts together an elegant way of reinvigorating this part of the metabolic theory of ecology. He uses quantile regressions to model just the upper parts of the relationship between species richness and mean annual temperature, rather than modelling its central tendency through the classical linear regression family of methods –as was done in the past. This assumes that the baseline effect of ambient energy does produce the negative linear relationship between richness and temperature predicted by the kinetic model (Allen et al. 2002), but also that this effect only poses an upper limit for species richness, and the effects of other factors may result in lower levels of species co-occurrence, thus producing a triangular rather than linear relationship. The results of Segovia’s simple and elegant analytical design show unequivocally that the predictions of the kinetic model become progressively more explanatory towards the upper quartiles of the relationship between species richness and temperature along over 10,000 tree local inventories throughout the Americas, reaching over 70% of explanatory power for the upper 5% of the relationship (i.e. the 95% quantile). This confirms to a large extent his reformulation of the predictions of the kinetic model.
Further, the neat study from Segovia (2021) also provides evidence confirming that the well-known spatial non-stationarity in the richness–temperature relationship (see Cassemiro et al. 2007) also applies to its upper-bound segment. Both the explanatory power and the slope of the relationship in the 95% upper quantile vary widely between biomes, reaching values similar to the predictions of the kinetic model only in cold temperate environments –precisely where temperature becomes more important than water availability as a constrain to plant life (O’Brien 1998; Hawkins et al. 2003). Part of these variations are indeed related with changes in water deficit and number of frost days along the XXth Century, as shown by the residuals of this paper (Segovia 2021) and a more detailed separate study (Segovia et al. 2020). This pinpoints the importance of the relative balance between water and energy as two of the main climatic factors constraining species diversity gradients, confirming the value of hypotheses that date back to Humboldt’s work (see Hawkins 2001, 2008). There is however a significant amount of unexplained variation in Segovia’s analyses, in particular in the progressive departure of the predictions of the kinetic model as we move towards the tropics, or downwards along the lower quantiles of the richness–temperature relationship. This calls for a deeper exploration of the factors that modify the baseline relationship between richness and energy, opening a new avenue for the macroecological investigation of how different forces and processes shape up geographical diversity gradients beyond the mere energetic constrains imposed by the basal limitations of multicellular life on Earth.
References
Algar, A.C., Kerr, J.T. and Currie, D.J. (2007) A test of Metabolic Theory as the mechanism underlying broad-scale species-richness gradients. Global Ecology and Biogeography, 16, 170-178. doi: https://doi.org/10.1111/j.1466-8238.2006.00275.x
Allen, A.P., Brown, J.H. and Gillooly, J.F. (2002) Global biodiversity, biochemical kinetics, and the energetic-equivalence rule. Science, 297, 1545-1548. doi: https://doi.org/10.1126/science.1072380
Brown, J.H., Gillooly, J.F., Allen, A.P., Savage, V.M. and West, G.B. (2004) Toward a metabolic theory of ecology. Ecology, 85, 1771-1789. doi: https://doi.org/10.1890/03-9000
Cassemiro, F.A.d.S., Barreto, B.d.S., Rangel, T.F.L.V.B. and Diniz-Filho, J.A.F. (2007) Non-stationarity, diversity gradients and the metabolic theory of ecology. Global Ecology and Biogeography, 16, 820-822. doi: https://doi.org/10.1111/j.1466-8238.2007.00332.x
Currie, D.J. (1991) Energy and large-scale patterns of animal- and plant-species richness. The American Naturalist, 137, 27-49. doi: https://doi.org/10.1086/285144
Currie, D.J., Mittelbach, G.G., Cornell, H.V., Field, R., Guegan, J.-F., Hawkins, B.A., Kaufman, D.M., Kerr, J.T., Oberdorff, T., O'Brien, E. and Turner, J.R.G. (2004) Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecology Letters, 7, 1121-1134. doi: https://doi.org/10.1111/j.1461-0248.2004.00671.x
D'Amen, M., Rahbek, C., Zimmermann, N.E. and Guisan, A. (2017) Spatial predictions at the community level: from current approaches to future frameworks. Biological Reviews, 92, 169-187. doi: https://doi.org/10.1111/brv.12222
Francis, A.P. and Currie, D.J. (2003) A globally consistent richness-climate relationship for Angiosperms. American Naturalist, 161, 523-536. doi: https://doi.org/10.1086/368223
Hawkins, B.A. (2001) Ecology's oldest pattern? Trends in Ecology & Evolution, 16, 470. doi: https://doi.org/10.1016/S0169-5347(01)02197-8
Hawkins, B.A. (2008) Recent progress toward understanding the global diversity gradient. IBS Newsletter, 6.1, 5-8. https://escholarship.org/uc/item/8sr2k1dd
Hawkins, B.A., Field, R., Cornell, H.V., Currie, D.J., Guégan, J.-F., Kaufman, D.M., Kerr, J.T., Mittelbach, G.G., Oberdorff, T., O'Brien, E., Porter, E.E. and Turner, J.R.G. (2003) Energy, water, and broad-scale geographic patterns of species richness. Ecology, 84, 3105-3117. doi: https://doi.org/10.1890/03-8006
Hawkins, B.A., Diniz-Filho, J.A.F., Jaramillo, C.A. and Soeller, S.A. (2006) Post-Eocene climate change, niche conservatism, and the latitudinal diversity gradient of New World birds. Journal of Biogeography, 33, 770-780. doi: https://doi.org/10.1111/j.1365-2699.2006.01452.x
Hawkins, B.A., Albuquerque, F.S., Araújo, M.B., Beck, J., Bini, L.M., Cabrero-Sañudo, F.J., Castro Parga, I., Diniz-Filho, J.A.F., Ferrer-Castán, D., Field, R., Gómez, J.F., Hortal, J., Kerr, J.T., Kitching, I.J., León-Cortés, J.L., et al. (2007a) A global evaluation of metabolic theory as an explanation for terrestrial species richness gradients. Ecology, 88, 1877-1888. doi:10.1890/06-1444.1. doi: https://doi.org/10.1890/06-1444.1
Hawkins, B.A., Diniz-Filho, J.A.F., Bini, L.M., Araújo, M.B., Field, R., Hortal, J., Kerr, J.T., Rahbek, C., Rodríguez, M.Á. and Sanders, N.J. (2007b) Metabolic theory and diversity gradients: Where do we go from here? Ecology, 88, 1898–1902. doi: https://doi.org/10.1890/06-2141.1
Hortal, J., Rodríguez, J., Nieto-Díaz, M. and Lobo, J.M. (2008) Regional and environmental effects on the species richness of mammal assemblages. Journal of Biogeography, 35, 1202–1214. doi: https://doi.org/10.1111/j.1365-2699.2007.01850.x
Kreft, H. and Jetz, W. (2007) Global patterns and determinants of vascular plant diversity. Proceedings of the National Academy of Sciences USA, 104, 5925-5930. doi: https://doi.org/10.1073/pnas.0608361104
O'Brien, E. (1998) Water-energy dynamics, climate, and prediction of woody plant species richness: an interim general model. Journal of Biogeography, 25, 379-398. doi: https://doi.org/10.1046/j.1365-2699.1998.252166.x
Ricklefs, R.E. (1987) Community diversity: Relative roles of local and regional processes. Science, 235, 167-171. doi: https://doi.org/10.1126/science.235.4785.167
Ricklefs, R.E. (2004) A comprehensive framework for global patterns in biodiversity. Ecology Letters, 7, 1-15. doi: https://doi.org/10.1046/j.1461-0248.2003.00554.x
Ricklefs, R.E. (2008) Disintegration of the ecological community. American Naturalist, 172, 741-750. doi: https://doi.org/10.1086/593002
Segovia, R.A. (2021) Temperature predicts the maximum tree-species richness and water and frost shape the residual variation. bioRxiv, 836338, ver. 4 peer-reviewed and recommended by Peer community in Ecology. doi: https://doi.org/10.1101/836338
Segovia, R.A., Pennington, R.T., Baker, T.R., Coelho de Souza, F., Neves, D.M., Davis, C.C., Armesto, J.J., Olivera-Filho, A.T. and Dexter, K.G. (2020) Freezing and water availability structure the evolutionary diversity of trees across the Americas. Science Advances, 6, eaaz5373. doi: https://doi.org/10.1126/sciadv.aaz5373
| Temperature predicts the maximum tree-species richness and water and frost shape the residual variation | Ricardo A. Segovia | <p>The kinetic hypothesis of biodiversity proposes that temperature is the main driver of variation in species richness, given its exponential effect on biological activity and, potentially, on rates of diversification. However, limited support fo... |  | Biodiversity, Biogeography, Botany, Macroecology, Species distributions | Joaquín Hortal | | 2019-11-10 20:56:40 | View |
Body temperatures, life history, and skeletal morphology in the nine-banded armadillo (Dasypus novemcinctus)
Frank Knight, Cristin Connor, Ramji Venkataramanan, Robert J. Asher
https://doi.org/10.17863/CAM.50971
Is vertebral count in mammals influenced by developmental temperature? A study with Dasypus novemcinctus
Recommended by Mar Sobral based on reviews by Darin Croft and ?
Mammals show a very low level of variation in vertebral count, both among and within species, in comparison to other vertebrates [1]. Jordan’s rule for fishes states that the vertebral number among species increases with latitude, due to ambient temperatures during development [2]. Temperature has also been shown to influence vertebral count within species in fish [3], amphibians [4], and birds [5]. However, in mammals the count appears to be constrained, on the one hand, by a possible relationship between the development of the skeleton and the proliferations of cell lines with associated costs (neural malformations, cancer etc., [6]), and on the other by the cervical origin of the diaphragm [7].
Knight et al. [8] investigate the effect of intrauterine temperature variation on skeletal morphology during development, and focus on a particular mammal, Dasypus novemcinctus, or nine-banded armadillo. Armadillos (Xenarthra) and are characterized by relatively low body temperatures and low basal rates of metabolism. Dasypus novemcinctus is the only xenarthran mammal to have naturally expanded its range into the middle latitudes of the U.S., and one of the few mammals that invaded North America from South America. It is one of few placentals that withstand considerable decrease of body temperature without torpor. It presents a resting body temperature that is low and variable for a placental mammal of its size [9] and is the only vertebrate that gives birth to monozygotic quadruplets. Among 42 monotreme, marsupial and placental genera, Dasypus novemcinctus shows the highest variation of thoracolumbar vertebral count [10].
The particularities of Dasypus novemcinctus regarding vertebral count variation and ability to withstand variable temperature qualify it as a target organism for study of the relationship between skeleton morphology and temperature in mammals.
Knight et al. [8] explored variability in vertebral count within Dasypus novemcinctus to understand whether temperature during development determines skeleton morphology. To this end they experimented with 22 armadillos (19 with data) and litters from 12 pregnant females, in two environments, for three years — an impressive effort and experimental setup. Moreover, they used a wide variety of advanced experimental and analytical techniques. For example, they implanted intra-abdominal, long-term temperature recorders, which recorded data every 6 to 120 minutes for up to several months. They analysed body temperature periodicity by approximation of the recordings with Fourier series, and they CT-scanned fetuses.
All 19 individuals (from which data could be gathered) exhibited substantial daily variation in body temperature. Several intriguing results emerged such as the counter-intuitive finding that the mammals’ body temperature fluctuates more indoors than outdoors. Furthermore, three females (out of 12) were found to have offspring with atypical skeletons, and two of these mothers presented an extremely low internal temperature early in pregnancy. Additionally, genetically identical quadruplets differed skeletally among themselves within two litters.
Results are not yet definitive about the relationship of temperature during development and vertebral count in Dasypus novemcinctus. However, Knight et al. [8] demonstrated that nine-banded armadillos survive with high daily internal temperature fluctuations and successfully bring to term offspring which vary in skeletal morphology among and within genetically identical litters despite major temperature extremes.
References
[1] Hautier L, Weisbecker V, Sánchez-Villagra MR, Goswami A, Asher RJ (2010) Skeletal development in sloths and the evolution of mammalian vertebral patterning. Proceedings of the National Academy of Sciences, 107, 18903–18908. doi: 10.1073/pnas.1010335107
[2] Jordan, D.S. (1892) Relations of temperature to vertebrae among fishes. Proceedings of the United States National Museum, 1891, 107-120. doi: 10.5479/si.00963801.14-845.107
[3] Tibblin P, Berggren H, Nordahl O, Larsson P, Forsman A (2016) Causes and consequences of intra-specific variation in vertebral number. Scientific Reports, 6, 1–12. doi: 10.1038/srep26372
[4] Peabody RB, Brodie ED (1975) Effect of temperature, salinity and photoperiod on the number of trunk vertebrae in Ambystoma maculatum. Copeia, 1975, 741–746. doi: 10.2307/1443326
[5] Lindsey CC, Moodie GEE (1967) The effect of incubation temperature on vertebral count in the chicken. Canadian Journal of Zoology, 45, 891–892. doi: 10.1139/z67-099
[6] Galis F, Dooren TJMV, Feuth JD, Metz JAJ, Witkam A, Ruinard S, Steigenga MJ, Wunaendts LCD (2006) Extreme selection in humans against homeotic transformations of cervical vertebrae. Evolution, 60, 2643–2654. doi: 10.1111/j.0014-3820.2006.tb01896.x
[7] Buchholtz EA, Stepien CC (2009) Anatomical transformation in mammals: developmental origin of aberrant cervical anatomy in tree sloths. Evolution and Development, 11, 69–79. doi: 10.1111/j.1525-142X.2008.00303.x
[8] Knight F, Connor C, Venkataramanan R, Asher RJ. (2020). Body temperatures, life history, and skeletal morphology in the nine-banded armadillo (Dasypus novemcinctus). PCI-Ecology. doi: 10.17863/CAM.50971
[9] McNab BK (1980) Energetics and the limits to a temperate distribution in armadillos. Journal of Mammalogy, 61, 606–627. doi: 10.2307/1380307
[10] Asher RJ, Lin KH, Kardjilov N, Hautier L (2011) Variability and constraint in the mammalian vertebral column. Journal of Evolutionary Biology, 24, 1080–1090. doi: 10.1111/j.1420-9101.2011.02240.x
| Body temperatures, life history, and skeletal morphology in the nine-banded armadillo (Dasypus novemcinctus) | Frank Knight, Cristin Connor, Ramji Venkataramanan, Robert J. Asher | <p>The nine banded armadillo (*Dasypus novemcinctus*) is the only xenarthran mammal to have naturally expanded its range into the middle latitudes of the USA. It is not known to hibernate, but has been associated with unusually labile core body te... | 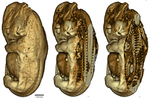 | Behaviour & Ethology, Evolutionary ecology, Life history, Physiology, Zoology | Mar Sobral | | 2019-11-22 22:57:31 | View |
Environmental variables determining the distribution of an avian parasite: the case of the Philornis torquans complex (Diptera: Muscidae) in South America
Pablo F. Cuervo, Alejandro Percara, Lucas Monje, Pablo M. Beldomenico, Martín A. Quiroga
https://doi.org/10.1101/839589
Catching the fly in dystopian times
Recommended by Rodrigo Medel based on reviews by 4 anonymous reviewers
Host-parasite interactions are ubiquitous on Earth. They are present in almost every conceivable ecosystem and often result from a long history of antagonist coevolution [1,2]. Recent studies on climate change have revealed, however, that modification of abiotic variables are often accompanied by shifts in the distributional range of parasites to habitats far beyond their original geographical distribution, creating new interactions in novel habitats with unpredictable consequences for host community structure and organization [3,4]. This situation may be especially critical for endangered host species having small population abundance and restricted distribution range. The infestation of bird species with larvae of the muscid fly genus Philornis is a case in point. At least 250 bird species inhabiting mostly Central and South America are infected by Philornis flies [5,6]. Fly larval development occurs in bird faeces, nesting material, or inside nestlings, affecting the development and nestling survival.
Recent reports indicate significant reduction of bird numbers associated with recent Philornis infection, the most conspicuous being Galapagos finches [7,8]. One way to prevent this potential effect consists in to examine the expected geographical shift of Philornis fly species under future climate change scenarios so that anticipatory conservation practices become implemented for endangered bird species. In this regard, Ecological Niche Modeling (ENM) techniques have been increasingly used as a useful tool to predict disease transmission as well as the species becoming infected under different climate change scenarios [9-11]. The paper of Cuervo et al. [12] is an important advance in this regard. By identifying for the first time the macro-environmental variables influencing the abiotic niche of species of the Philornis torquans complex in southern South America, the authors perform a geographical projection model that permits identification of the areas susceptible to be colonized by Philornis species in Argentina, Brazil, and Chile, including habitats where the parasitic fly is still largely absent at present. Their results are promissory for conservation studies and contribute to the still underdeveloped issue of the way climate change impacts on antagonistic ecological relationships.
References
[1] Thompson JN (1994) The Coevolutionary Process. University of Chicago Press.
[2] Poulin R (2007) Evolutionary Ecology of Parasites: (Second Edition). Princeton University Press. doi: 10.2307/j.ctt7sn0x
[3] Pickles RSA, Thornton D, Feldman R, Marques A, Murray DL (2013) Predicting shifts in parasite distribution with climate change: a multitrophic level approach. Global Change Biology, 19, 2645–2654. doi: 10.1111/gcb.12255
[4] Marcogliese DJ (2016) The distribution and abundance of parasites in aquatic ecosystems in a changing climate: More than just temperature. Integrative and Comparative Biology, 56, 611–619. doi: 10.1093/icb/icw036
[5] Dudaniec RY, Kleindorfer S (2006) Effects of the parasitic flies of the genus Philornis (Diptera: Muscidae) on birds. Emu - Austral Ornithology, 106, 13–20. doi: 10.1071/MU04040
[6] Antoniazzi LR, Manzoli DE, Rohrmann D, Saravia MJ, Silvestri L, Beldomenico PM (2011) Climate variability affects the impact of parasitic flies on Argentinean forest birds. Journal of Zoology, 283, 126–134. doi: 10.1111/j.1469-7998.2010.00753.x
[7] Fessl B, Sinclair BJ, Kleindorfer S (2006) The life-cycle of Philornis downsi (Diptera: Muscidae) parasitizing Darwin’s finches and its impacts on nestling survival. Parasitology, 133, 739–747. doi: 10.1017/S0031182006001089
[8] Kleindorfer S, Peters KJ, Custance G, Dudaniec RY, O’Connor JA (2014) Changes in Philornis infestation behavior threaten Darwin’s finch survival. Current Zoology, 60, 542–550. doi: 10.1093/czoolo/60.4.542
[9] Johnson EE, Escobar LE, Zambrana-Torrelio C (2019) An ecological framework for modeling the geography of disease transmission. Trends in Ecology and Evolution, 34, 655–668. doi: 10.1016/j.tree.2019.03.004
[10] Carvalho BM, Rangel EF, Ready PD, Vale MM (2015) Ecological niche modelling predicts southward expansion of Lutzomyia (Nyssomyia) flaviscutellata (Diptera: Psychodidae: Phlebotominae), vector of Leishmania (Leishmania) amazonensis in South America, under climate change. PLOS ONE, 10, e0143282. doi: 10.1371/journal.pone.0143282
[11] Garrido R, Bacigalupo A, Peña-Gómez F, Bustamante RO, Cattan PE, Gorla DE, Botto-Mahan C (2019) Potential impact of climate change on the geographical distribution of two wild vectors of Chagas disease in Chile: Mepraia spinolai and Mepraia gajardoi. Parasites and Vectors, 12, 478. doi: 10.1186/s13071-019-3744-9
[12] Cuervo PF, Percara A, Monje L, Beldomenico PM, Quiroga MA (2020) Environmental variables determining the distribution of an avian parasite: the case of the Philornis torquans complex (Diptera: Muscidae) in South America. bioRxiv, 839589, ver. 5 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/839589
| Environmental variables determining the distribution of an avian parasite: the case of the Philornis torquans complex (Diptera: Muscidae) in South America | Pablo F. Cuervo, Alejandro Percara, Lucas Monje, Pablo M. Beldomenico, Martín A. Quiroga | <p>*Philornis* flies are the major cause of myasis in altricial nestlings of neotropical birds. Its impact ranges from subtle to lethal, being of major concern in endangered bird species with geographically-restricted, fragmented and small-sized p... | 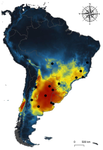 | Biogeography, Macroecology, Parasitology, Species distributions | Rodrigo Medel | | 2019-11-26 21:31:33 | View |
Trophic cascade driven by behavioural fine-tuning as naïve prey rapidly adjust to a novel predator
Chris J Jolly, Adam S Smart, John Moreen, Jonathan K Webb, Graeme R Gillespie and Ben L Phillips
https://doi.org/10.1101/856997
While the quoll’s away, the mice will play… and the seeds will pay
Recommended by Denis Réale based on reviews by 2 anonymous reviewers
A predator can strongly influence the demography of its prey, which can have profound carryover effects on the trophic network; so-called density-mediated indirect interactions (DMII; Werner and Peacor 2003; Schmitz et al. 2004; Trussell et al. 2006). Furthermore, a novel predator can alter the phenotypes of its prey for traits that will change prey foraging efficiency. These trait-mediated indirect interactions may in turn have cascading effects on the demography and features of the basal resources consumed by the intermediate consumer (TMIII; Werner and Peacor 2003; Schmitz et al. 2004; Trussell et al. 2006), but very few studies have looked for these effects (Trusell et al. 2006). The study “Trophic cascade driven by behavioural fine-tuning as naïve prey rapidly adjust to a novel predator”, by Jolly et al. (2020) is therefore a much-needed addition to knowledge in this field. The authors have profited from a rare introduction of Northern quolls (Dasyurus hallucatus) on an Australian island, to examine both the density-mediated and trait-mediated indirect interactions with grassland melomys (Melomys burtoni) and the vegetation of their woodland habitat.
Jolly et al. (2020) compared melomys populations in four quoll-invaded and three quoll-free sites on the same island. Using capture-mark-recapture methods, they found a lower survival and decreased population size in quoll-invaded sites compared to quoll-free sites. Although they acknowledge that this decline could be attributable to either the direct effects of the predator or to a wildfire that occurred early in the experiment in the quoll-invaded sites, the authors argue that the wildfire alone cannot explain all of their results.
Beyond demographic effects, Jolly et al. (2020) also examined risk taking, foraging behaviour, and predator avoidance in melomys. Quoll presence was first associated with a strong decrease in risk taking in melomys, but the difference disappeared over the three years of study, indicating a possible adjustment by the prey. In quoll-invaded sites, though, melomys continued to be more neophobic than in the quoll-free sites throughout the study. Furthermore, in a seed (i.e. wheat) removal experiment, Jolly et al. (2020) measured how melomys harvested seeds in the presence or absence of predator scents. In both quoll-invaded and quoll-free sites, melomys density increased seed harvest efficiency. Melomys also removed less seeds in quoll-invaded sites than in quoll-free sites, supporting both the DMII and TMII hypotheses. However, in the quoll-invaded sites only, melomys foraged less on predator-scented seed patches than on unscented ones, trading foraging efficiency for an increased safety against predators, and this effect increased across the years. This last result indicates that predators can indirectly influence seed consumption through the trade-off between foraging and predator avoidance, strongly supporting the TMII hypothesis.
Ideally, the authors would have run a nice before-after, impact-control design, but nature does not always allow for ideal experimental designs. Regardless, the results of such an “experiment in the wild” predation study are still valuable, as they are very rare (Trussell et al. 2006), and they provide crucial information on the direct and indirect interactions along a trophic cascade. Furthermore, the authors have effectively addressed any concerns about potential confounding factors, and thus have a convincing argument that their results represent predator-driven demographic and behavioural changes.
One important question remains from an evolutionary ecology standpoint: do the responses of melomys to the presence of quolls represent phenotypically plastic changes or rapid evolutionary changes caused by novel selection pressures? Classically, TMII are assumed to be mostly caused by phenotypic plasticity (Werner and Peacor 2003), and this might be the case when the presence of the predator is historical. Phenotypic plasticity allows quick and reversible adjustments of the prey population to changes in the predator density. When the predator population declines, such rapid phenotypic changes can be reversed, reducing the cost associated with anti-predator behaviour (e.g., lower foraging efficiency) in the absence of predators. In the case of a novel predator, however, short-term evolutionary responses by the prey may play role in the TMII, as they would allow a phenotypic shift in prey’s traits along the trade-off between foraging efficiency and anti-predator response that will probably more advantageous over the longer term, if the predator does not disappear. The authors state that they could not rule out one or the other of these hypotheses. However, future work estimating the relative importance of phenotypic plasticity and evolutionary changes in the quoll-melomys system would be valuable. Phenotypic selection analysis, for example, by estimating the link between survival and the traits measured, might help test for a fitness advantage to altered behaviour in the presence of a predator. Common garden experiments, comparing the quoll-invaded and the quoll-free melomys populations, might also provide information on any potential evolutionary changes caused by predation. More work could also analyse the potential effects on the seed populations. Not only might the reduction in seed predation have consequences on the landscape in the future, as the authors mention, but it may also mean that the seeds themselves could be subject to novel selection pressures, which may affect their phenology, physiology or life history. Off course, the authors will have to switch from wheat to a more natural situation, and evaluate the effects of changes in the melomys population on the feature of the local vegetation and the ecosystem.
Finally, the authors have not yet found that the observed changes in the traits have translated into a demographic rebound for melomys. Here again, I can see an interesting potential for further studies. Should we really expect an evolutionary rescue (Bell and Gonzalez 2009) in this system? Alternatively, should the changes in behaviour be accompanied by permanent changes in life history, such as a slower pace-of-life (Réale et al. 2010) that could possibly lead to lower melomys density?
This paper provides nice in natura evidence for density- and trait-mediated indirect interactions hypotheses. I hope it will be the first of a long series of work on this interesting quoll-melomys system, and that the authors will be able to provide more information on the eco-evolutionary consequences of a novel predator on a trophic network.
References
-Bell G, Gonzalez A (2009) Evolutionary rescue can prevent extinction following environmental change. Ecology letters, 12(9), 942-948. https://doi.org/10.1111/j.1461-0248.2009.01350.x
-Jolly CJ, Smart AS, Moreen J, Webb JK, Gillespie GR, Phillips BL (2020) Trophic cascade driven by behavioural fine-tuning as naïve prey rapidly adjust to a novel predator. bioRxiv, 856997, ver. 6 peer-reviewed and recommended by PCI Ecology. https://doi.org/ 10.1101/856997
-Matassa C, Ewanchuk P, Trussell G (2018) Cascading effects of a top predator on intraspecific competition at intermediate and basal trophic levels. Functional Ecology, 32(9), 2241-2252. https://doi.org/10.1111/1365-2435.13131
-Réale D, Garant D, Humphries MM, Bergeron P, Careau V, Montiglio PO (2010) Personality and the emergence of the pace-of-life syndrome concept at the population level. Philosophical Transactions of the Royal Society B: Biological Sciences, 365(1560), 4051-4063. https://doi.org/10.1098/rstb.2010.0208
-Schmitz O, Krivan V, Ovadia O (2004) Trophic cascades: the primacy of trait‐mediated indirect interactions. Ecology Letters 7(2), 153-163. https://doi.org/10.1111/j.1461-0248.2003.00560.x
-Trussell G, Ewanchuk P, Matassa C (2006). Habitat effects on the relative importance of trait‐ and density‐mediated indirect interactions. Ecology Letters, 9(11), 1245-1252. https://doi.org/10.1111/j.1461-0248.2006.00981.x
-Werner EE, Peacor SD (2003) A review of trait‐mediated indirect interactions in ecological communities. Ecology, 84(5), 1083-1100. https://doi.org/10.1890/0012-9658(2003)084[1083:AROTII]2.0.CO;2
| Trophic cascade driven by behavioural fine-tuning as naïve prey rapidly adjust to a novel predator | Chris J Jolly, Adam S Smart, John Moreen, Jonathan K Webb, Graeme R Gillespie and Ben L Phillips | <p>The arrival of novel predators can trigger trophic cascades driven by shifts in prey numbers. Predators also elicit behavioural change in prey populations, via phenotypic plasticity and/or rapid evolution, and such changes may also contribute t... |  | Behaviour & Ethology, Biological invasions, Evolutionary ecology, Experimental ecology, Foraging, Herbivory, Population ecology, Terrestrial ecology, Tropical ecology | Denis Réale | | 2019-11-27 21:39:44 | View |
Studying a rare behavior in a polygamous bird: male parental care in great-tailed grackles
Recommended by Marie-Jeanne Holveck based on reviews by Matthieu Paquet and André C Ferreira
The Great-tailed grackle (Quiscalus mexicanus) is a polygamous bird species that is originating from Central America and rapidly expanding its geographic range toward the North, and in which females were long thought to be the sole nest builders and caretakers of the young. In their pre-registration [1], Folsom and collaborators report repeated occurrences of male parental care and develop hypotheses aiming at better understanding the occurrence and the fitness consequences of this very rarely observed male behavior. They propose to assess if male parental care correlates with the circulating levels of several relevant hormones, increases offspring survival, is a local adaptation, and is a mating strategy, in surveying three populations located in Arizona (middle of the geographic range expansion), California (northern edge of the geographic range), and in Central America (core of the range). This study is part of a 5-year bigger project.
Both reviewers and I strongly value Folsom and collaborators’ commitment to program a study, in natural field conditions, of a rare, yet likely evolutionary-important behavior, namely parental care by males of the great-tailed grackle. Yet, we all also recognized that it is a risky endeavor, and as a consequence, we wondered about the authors’ ability to reach a sufficient sample size to statistically test (all) hypotheses and predictions with enough confidence (e.g. risk of type I errors, also known as false positives).
Folsom and collaborators acknowledged these limitations in their pre-registration. (i) They made the exploratory nature of their research work clear to readers. (ii) They adapted their analysis plan in running prior power analyses and in focusing on effect sizes (estimates and confidence intervals). (iii) Last and not least, Folsom and collaborators clearly exposed a priori hypotheses, their associated predictions and alternatives, and ranked the latter based on their plausibility according to knowledge in the current and other study systems. Developing theory about male parental care behavior more generally with regard to a polygamous species that is rapidly expanding its geographic range and that is considered not to provide male parental care is without any doubt an added value to this study.
In summary, while this study will likely be insufficient to fully understand male parental care behavior of great-tailed grackles, it is much needed because it will definitely allow rejecting some predictions (e.g., if this behavior is present in all the studied populations, it would be common across range against expectation; finding only one male providing care to an unrelated offspring would lead to reject the prediction that males only care for their own offspring) and thus it will help laying the foundation of future research directions.
I strongly support the pre-registration system and thank all the contributors for producing a fruitful discussion throughout the review process, which in fine improved the clarity and logic of this pre-registration. Given the positive and encouraging reviews, the detailed and thorough answers to all comments by Folsom and collaborators, and their satisfactory and interesting revisions, I am happy to recommend this pre-registration and I look forward to seeing its outcomes.
References
[1] Folsom MA, MacPherson M, Lukas D, McCune KB, Bergeron L, Bond A, Blackwell A, Rowney C, Logan CJ. 2020. Investigating the rare behavior of male parental care in great-tailed grackles. corinalogan.com/Preregistrations/gmalecare.html In principle acceptance by PCI Ecology of the version on 15 June 2020 corinalogan/grackles/blob/master/Files/Preregistrations/gmalecare.Rmd.
| Investigating the rare behavior of male parental care in great-tailed grackles | Folsom MA, MacPherson M, Lukas D, McCune KB, Bergeron L, Bond A, Blackwell A, Rowney C, Logan CJ | This is a PREREGISTRATION submitted for pre-study peer review. Our planned data collection START DATE is May 2020, therefore it would be ideal if the peer review process could be completed before then.
Abstract: Great-tailed grackles (Quiscalus... |  | Behaviour & Ethology, Biological invasions, Preregistrations, Zoology | Marie-Jeanne Holveck | | 2019-12-05 17:38:47 | View |







