Touchy matter: the delicate balance between Morgan’s canon and open-minded description of advanced cognitive skills in the animal
Recommended by Francois-Xavier Dechaume-Moncharmont based on reviews by Valérie Dufour and Alex Taylor based on reviews by Valérie Dufour and Alex Taylor
In a recent paper published in PNAS, Fayet et al. [1] reported scarce field observations of two Atlantic puffins (four years apart) apparently scratching their bodies using sticks, which was interpreted by the authors as evidence of tool use in this species. In a short response, Benjamin Farrar [2] raises serious concerns about this interpretation and proposes simpler, more parsimonious, mechanisms explaining the observed behaviour: a textbook case of Morgan's canon.
In virtually all introductory lectures on animal behaviour, students are advised to exercise caution when interpreting empirical data and weighting alternative explanations. We are sometimes prisoner of our assumptions: our desire of beliefs in advanced cognitive skills in non-human species make us more receptive to facts confirming our preconceptions than to simpler, less exciting, interpretations (a phenomenon known as "confirmation bias" in psychology). We must resist the temptation to accept appealing explanations without enough critical thinking. Our students are thus taught to apply the Lloyd Morgan's canon, a variant of one of the most important heuristics in Science, the principle of parsimony or Occam's razor, rephrased by Morgan [3, page 53] in the context of animal behaviour: "In no case may we interpret an action as the outcome of a higher psychical faculty, if it can be interpreted as the outcome of the exercise of one that stands lower in the psychological scale". In absence of evidence to the contrary, one should postulate the simplest cognitive skill consistent with the observed behaviour. While sometimes criticized from an epistemological point of view [4-6], it remains an essential and largely accepted framework of animal cognition. It has repeatedly proved to be a useful guide in the minefield of comparative psychology. Classical ethology questions related to the existence of, for instance, meta-cognition [7], intentionality or problem solving [8] have been convincingly investigated using this principle.
Yet, there is a downside to this conservative approach. Blind reference to Morgan's canon may narrow our theoretical thinking about animal cognition [7,9]. It could be counter-productive to systematically deny advanced cognitive skills in animals. On the contrary, keeping our mind open to unplanned observations, unexpected discoveries, or serendipity [10], and being prepared to accept new hypotheses, sometimes fairly remote from the dominant paradigm, may be a fruitful research strategy. To quote Darwin's famous letter to Alfred Wallace: "I am a firm believer, that without speculation there is no good and original observation" [11]. Brief notes in specialized scientific journals, or even in grey literature (by enthusiast amateur ornithologists, ichthyologists, or entomologists), constitutes a rich array of anecdotal observations. For instance, Sol et al. [12] convincingly compared the innovation propensity across bird species by screening ornithology literature using keywords like 'never reported', 'not seen before', 'first report', 'unusual' or 'novel'. Even if "the plural of anecdote is not data" as the saying goes, such descriptions of novel behaviours, even single-subject observations, are indisputably precious: taxonomic ubiquity of a behaviour is a powerful argument in favour of evolutionary convergence. Of course, a race to the bottom, amplified by the inevitable media hypes around scientific articles questioning human exceptionalism, is another possible scientific trap for behavioural biologists in search of skills characteristic of so-called advanced species, but never described so far in supposedly cognitively simpler organisms. As stated by Franz de Waal [9]: "I have nothing against anecdotes, especially if they have been caught on camera or come from reputable observers who know their animals; but I do view them as a starting point of research, never an end point".
In the case of the two video observations of puffins apparently using sticks as scratching tool, it must be considered as a mere anecdote unless scientists systematically investigate this behaviour. In his constructive criticism of Fayet et al.'s paper, Benjamin Farrar [2] proposes interesting directions of research and testable predictions. A correlation between the background rate of stick picking and the rate of stick preening would indicate that this behaviour was more likely explained by fluke than genuine innovation in this species.
References
[1] Fayet, A. L., Hansen, E. S., and Biro, D. (2020). Evidence of tool use in a seabird. Proceedings of the National Academy of Sciences, 117(3), 1277–1279. doi: 10.1073/pnas.1918060117
[2] Farrar, B. G. (2020). Evidence of tool use in a seabird? PsyArXiv, 463hk, ver. 5 recommended and peer-reviewed by Peer Community In Ecology. doi: 10.31234/osf.io/463hk
[3] Morgan, C. L. (1894). An introduction to comparative psychology. London, UK: Walter Scott, Ltd. Retrieved from https://archive.org/details/introductiontoco00morg/page/53/mode/2up
[4] Meketa, I. (2014). A critique of the principle of cognitive simplicity in comparative cognition. Biology and Philosophy, 29(5), 731–745. doi: 10.1007/s10539-014-9429-z
[5] Fitzpatrick, S. (2017). Against Morgan's Canon. In K. Andrews and J. Beck (Eds.), The Routledge handbook of philosophy of animal minds (pp. 437–447). London, UK: Routledge, Taylor and Francis Group. doi: 10.4324/9781315742250.ch42
[6] Starzak, T. (2017). Interpretations without justification: a general argument against Morgan's Canon. Synthese, 194(5), 1681–1701. doi: 10.1007/s11229-016-1013-4
[7] Arbilly, M., and Lotem, A. (2017). Constructive anthropomorphism: a functional evolutionary approach to the study of human-like cognitive mechanisms in animals. Proceedings of the Royal Society B: Biological Sciences, 284(1865), 20171616. doi: 10.1098/rspb.2017.1616
[8] Taylor, A. H., Knaebe, B., and Gray, R. D. (2012). An end to insight? New Caledonian crows can spontaneously solve problems without planning their actions. Proceedings of the Royal Society B: Biological Sciences, 279(1749), 4977–4981. doi: 10.1098/rspb.2012.1998
[9] de Waal, F. (2016). Are we smart enough to know how smart animals are? New-York, USA: W. W. Norton and Company.
[10] Scheffer, M. (2014). The forgotten half of scientific thinking. Proceedings of the National Academy of Sciences, 111(17), 6119–6119. doi: 10.1073/pnas.1404649111
[11] Darwin, C. R. (1857). Letter to A. R. Wallace, 22 December 1857. Retrieved 30 January 2020, from https://www.darwinproject.ac.uk/letter/DCP-LETT-2192.xml
[12] Sol, D., Lefebvre, L., and Rodríguez-Teijeiro, J. D. (2005). Brain size, innovative propensity and migratory behaviour in temperate Palaearctic birds. Proceedings of the Royal Society B: Biological Sciences, 272(1571), 1433–1441. doi: 10.1098/rspb.2005.3099
| Evidence of tool use in a seabird? | Benjamin G. Farrar | Fayet, Hansen and Biro (1) provide two observations of Atlantic puffins, *Fratercula arctica*, performing self-directed actions while holding a stick in their beaks. The authors interpret this as evidence of tool use as they suggest that the stick... |  | Behaviour & Ethology | Francois-Xavier Dechaume-Moncharmont | | 2020-01-22 11:55:27 | View |
Clumpy coexistence in phytoplankton: The role of functional similarity in community assembly
Caio Graco-Roza, Angel M. Segura, Carla Kruk, Patricia Domingos, Janne Soininen, Marcelo M. Marinho
https://doi.org/10.1101/869966
Environmental heterogeneity drives phytoplankton community assembly patterns in a tropical riverine system
Recommended by Cédric Hubas and Eric Goberville based on reviews by Eric Goberville and Dominique Lamy
What predisposes two individuals to form and maintain a relationship is a fundamental question. Using facial recognition to see whether couples' faces change over time to become more and more similar, psychology researchers have concluded that couples tend to be formed from the start between people whose faces are more similar than average [1]. As the saying goes, birds of a feather flock together.
And what about in nature? Are these rules of assembly valid for communities of different species?
In his seminal contribution, Robert MacArthur (1984) wrote ‘To do science is to search for repeated patterns’ [2]. Identifying the mechanisms that govern the arrangement of life is a hot research topic in the field of ecology for decades, and an absolutely essential prerequisite to answer the outstanding question of what shape ecological patterns in multi-species communities such as species-area relationships, relative species abundances, or spatial and temporal turnover of community composition; amid others [3]. To explain ecological patterns in nature, some rely on the concept that every species - through evolutionary processes and the acquisition of a unique set of traits that allow a species to be adapted to its abiotic and biotic environment - occupies a unique niche: Species coexistence comes as the result of niche differentiation [4,5]. Such a view has been challenged by the recognition of the key role of neutral processes [6], however, in which demographic stochasticity contributes to shape multi-species communities and to explain why congener species coexist much more frequently than expected by chance [7,8]. While the niche-based and neutral theories appear seemingly opposed at first sight [9], the dichotomy may be more philosophical than empirical [4,5]. Many examples have come to support that both concepts are not incompatible as they together influence the structure, diversity and functioning of communities [10], and are simply extreme cases of a continuum [11]. From this perspective, extrinsic factors, i.e., environmental heterogeneity, may influence the location of a given community along the niche-neutrality continuum.
The walk of species in nature is therefore neither random nor ecologically predestined. In microbial assemblages, the co-existence of these two antagonistic mechanisms has been shown both theoretically and empirically. It has been shown that a combination of stabilising (niche) and equalising (neutral) mechanisms was responsible for the existence of groups of coexistent species (clumps) in a phytoplankton rich community [12]. Analysing interannual changes (2003-2009) in the weekly abundance of diatoms and dinoflagellates located in a temperate coastal ecosystem of the Western English Channel, Mutshinda et al. [13] found a mixture of biomass dynamics consistent with the neutrality-niche continuum hypothesis. While niche processes explained the dynamic of phytoplankton functional groups (i.e., diatoms vs. dinoflagellates) in terms of biomass, neutral processes mainly dominated - 50 to 75% of the time - the dynamics at the species level within functional groups [13]. From one endpoint to another, defining the location of a community along the continuum is all matter of scale [4,11].
In their study, testing predictions made by an emergent neutrality model, Graco-Roza et al. [14] provide empirical evidence that neutral and niche processes joined together to shape and drive planktonic communities in a riverine ecosystem. Body size - the 'master trait' - is used here as a discriminant ecological dimension along the niche axis. From their analysis, they not only show that the specific abundance is organised in clumps and gaps along the niche axis, but also reveal that different clumps exist along the river course. They identify two main clumps in body size - with species belonging to three different morphologically-based functional groups - and characterise that among-species differences in biovolume are driven by functional redundancy at the clump level; species functional distinctiveness being related to the relative biovolume of species. By grouping their variables according to seasons (cold-dry vs. warm-wet) or river elevation profile (upper, medium and lower course), they hereby highlight how environmental heterogeneity contributes to shape species assemblages and their dynamics and conclude that emergent neutrality models are a powerful approach to explain species coexistence; and therefore ecological patterns.
References
[1] Tea-makorn PP, Kosinski M (2020) Spouses’ faces are similar but do not become more similar with time. Scientific Reports, 10, 17001. https://doi.org/10.1038/s41598-020-73971-8.
[2] MacArthur RH (1984) Geographical Ecology: Patterns in the Distribution of Species. Princeton University Press.
[3] Vellend M (2020) The Theory of Ecological Communities (MPB-57). Princeton University Press.
[4] Wennekes PL, Rosindell J, Etienne RS (2012) The Neutral—Niche Debate: A Philosophical Perspective. Acta Biotheoretica, 60, 257–271. https://doi.org/10.1007/s10441-012-9144-6.
[5] Gravel D, Guichard F, Hochberg ME (2011) Species coexistence in a variable world. Ecology Letters, 14, 828–839. https://doi.org/10.1111/j.1461-0248.2011.01643.x.
[6] Hubbell SP (2001) The Unified Neutral Theory of Biodiversity and Biogeography (MPB-32). Princeton University Press.
[7] Leibold MA, McPeek MA (2006) Coexistence of the Niche and Neutral Perspectives in Community Ecology. Ecology, 87, 1399–1410. https://doi.org/10.1890/0012-9658(2006)87[1399:COTNAN]2.0.CO;2.
[8] Pielou EC (1977) The Latitudinal Spans of Seaweed Species and Their Patterns of Overlap. Journal of Biogeography, 4, 299–311. https://doi.org/10.2307/3038189.
[9] Holt RD (2006) Emergent neutrality. Trends in Ecology & Evolution, 21, 531–533. https://doi.org/10.1016/j.tree.2006.08.003.
[10] Scheffer M, Nes EH van (2006) Self-organized similarity, the evolutionary emergence of groups of similar species. Proceedings of the National Academy of Sciences, 103, 6230–6235. https://doi.org/10.1073/pnas.0508024103.
[11] Gravel D, Canham CD, Beaudet M, Messier C (2006) Reconciling niche and neutrality: the continuum hypothesis. Ecology Letters, 9, 399–409. https://doi.org/10.1111/j.1461-0248.2006.00884.x.
[12] Vergnon R, Dulvy NK, Freckleton RP (2009) Niches versus neutrality: uncovering the drivers of diversity in a species-rich community. Ecology Letters, 12, 1079–1090. https://doi.org/10.1111/j.1461-0248.2009.01364.x.
[13] Mutshinda CM, Finkel ZV, Widdicombe CE, Irwin AJ (2016) Ecological equivalence of species within phytoplankton functional groups. Functional Ecology, 30, 1714–1722. https://doi.org/10.1111/1365-2435.12641.
[14] Graco-Roza C, Segura AM, Kruk C, Domingos P, Soininen J, Marinho MM (2021) Clumpy coexistence in phytoplankton: The role of functional similarity in community assembly. bioRxiv, 869966, ver. 6 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/869966
| Clumpy coexistence in phytoplankton: The role of functional similarity in community assembly | Caio Graco-Roza, Angel M. Segura, Carla Kruk, Patricia Domingos, Janne Soininen, Marcelo M. Marinho | <p style="text-align: justify;">Emergent neutrality (EN) suggests that species must be sufficiently similar or sufficiently different in their niches to avoid interspecific competition. Such a scenario results in a transient pattern with clumps an... |  | Coexistence, Community ecology, Theoretical ecology | Cédric Hubas | | 2020-01-23 16:11:32 | View |
Pathogen community composition and co-infection patterns in a wild community of rodents
Jessica Lee Abbate, Maxime Galan, Maria Razzauti, Tarja Sironen, Liina Voutilainen, Heikki Henttonen, Patrick Gasqui, Jean-François Cosson, Nathalie Charbonnel
https://doi.org/10.1101/2020.02.09.940494
Reservoirs of pestilence: what pathogen and rodent community analyses can tell us about transmission risk
Recommended by Francois Massol based on reviews by Adrian Diaz, Romain Pigeault and 1 anonymous reviewer
Rodents are well known as one of the main animal groups responsible for human-transmitted pathogens. As such, it seems logical to try and survey what kinds of pathogenic microbes might be harboured by wild rodents, in order to establish some baseline surveillance and prevent future zoonotic outbreaks (Bernstein et al., 2022). This is exactly what Abbate et al. (2023) endeavoured and their findings are intimidating. Based on quite a large sampling effort, they collected more than 700 rodents of seven species around two villages in northeastern France. They looked for molecular markers indicative of viral and bacterial infections and proceeded to analyze their pathogen communities using multivariate techniques.
Variation in the prevalence of the different pathogens was found among host species, with e.g. signs of CPXV more prevalent in Cricetidae while some Mycoplasma strains were more prevalent in Muridae. Co-circulation of pathogens was found in all species, with some evidencing signs of up to 12 different pathogen taxa. The diversity of co-circulating pathogens was markedly different between host species and higher in adult hosts, but not affected by sex. The dataset also evinced some slight differences between habitats, with meadows harbouring a little more diversity of rodent pathogens than forests. Less intuitively, some pathogen associations seemed quite repeatable, such as the positive association of Bartonella spp. with CPXV in the montane water vole. The study allowed the authors to test several associations already described in the literature, including associations between different hemotropic Mycoplasma species.
I strongly invite colleagues interested in zoonoses, emerging pandemics and more generally One Health to read the paper of Abbate et al. (2023) and try to replicate them across the world. To prevent the next sanitary crises, monitoring rodents, and more generally vertebrates, population demographics is a necessary and enlightening step (Johnson et al., 2020), but insufficient. Following the lead of colleagues working on rodent ectoparasites (Krasnov et al., 2014), we need more surveys like the one described by Abbate et al. (2023) to understand the importance of the dilution effect in the prevalence and transmission of microbial pathogens (Andreazzi et al., 2023) and the formation of epidemics. We also need other similar studies to assess the potential of different rodent species to carry pathogens more or less capable of infecting other mammalian species (Morand et al., 2015), in other places in the world.
References
Abbate, J. L., Galan, M., Razzauti, M., Sironen, T., Voutilainen, L., Henttonen, H., Gasqui, P., Cosson, J.-F. & Charbonnel, N. (2023) Pathogen community composition and co-infection patterns in a wild community of rodents. BioRxiv, ver.4 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2020.02.09.940494
Andreazzi, C. S., Martinez-Vaquero, L. A., Winck, G. R., Cardoso, T. S., Teixeira, B. R., Xavier, S. C. C., Gentile, R., Jansen, A. M. & D'Andrea, P. S. (2023) Vegetation cover and biodiversity reduce parasite infection in wild hosts across ecological levels and scales. Ecography, 2023, e06579.
https://doi.org/10.1111/ecog.06579
Bernstein, A. S., Ando, A. W., Loch-Temzelides, T., Vale, M. M., Li, B. V., Li, H., Busch, J., Chapman, C. A., Kinnaird, M., Nowak, K., Castro, M. C., Zambrana-Torrelio, C., Ahumada, J. A., Xiao, L., Roehrdanz, P., Kaufman, L., Hannah, L., Daszak, P., Pimm, S. L. & Dobson, A. P. (2022) The costs and benefits of primary prevention of zoonotic pandemics. Science Advances, 8, eabl4183.
https://doi.org/10.1126/sciadv.abl4183
Johnson, C. K., Hitchens, P. L., Pandit, P. S., Rushmore, J., Evans, T. S., Young, C. C. W. & Doyle, M. M. (2020) Global shifts in mammalian population trends reveal key predictors of virus spillover risk. Proceedings of the Royal Society B: Biological Sciences, 287, 20192736.
https://doi.org/10.1098/rspb.2019.2736
Krasnov, B. R., Pilosof, S., Stanko, M., Morand, S., Korallo-Vinarskaya, N. P., Vinarski, M. V. & Poulin, R. (2014) Co-occurrence and phylogenetic distance in communities of mammalian ectoparasites: limiting similarity versus environmental filtering. Oikos, 123, 63-70.
https://doi.org/10.1111/j.1600-0706.2013.00646.x
Morand, S., Bordes, F., Chen, H.-W., Claude, J., Cosson, J.-F., Galan, M., Czirjak, G. Á., Greenwood, A. D., Latinne, A., Michaux, J. & Ribas, A. (2015) Global parasite and Rattus rodent invasions: The consequences for rodent-borne diseases. Integrative Zoology, 10, 409-423.
https://doi.org/10.1111/1749-4877.12143
| Pathogen community composition and co-infection patterns in a wild community of rodents | Jessica Lee Abbate, Maxime Galan, Maria Razzauti, Tarja Sironen, Liina Voutilainen, Heikki Henttonen, Patrick Gasqui, Jean-François Cosson, Nathalie Charbonnel | <p style="text-align: justify;">Rodents are major reservoirs of pathogens that can cause disease in humans and livestock. It is therefore important to know what pathogens naturally circulate in rodent populations, and to understand the factors tha... |  | Biodiversity, Coexistence, Community ecology, Eco-immunology & Immunity, Epidemiology, Host-parasite interactions, Population ecology, Species distributions | Francois Massol | | 2020-02-11 12:42:28 | View |
Hough transform implementation to evaluate the morphological variability of the moon jellyfish (Aurelia spp.)
Céline Lacaux, Agnès Desolneux, Justine Gadreaud, Bertrand Martin-Garin and Alain Thiéry
https://doi.org/10.1101/2020.03.11.986984
A new member of the morphometrics jungle to better monitor vulnerable lagoons
Recommended by Vincent Bonhomme based on reviews by Julien Claude and 1 anonymous reviewer
In the recent years, morphometrics, the quantitative description of shape and its covariation [1] gained considerable momentum in evolutionary ecology. Using the form of organisms to describe, classify and try to understand their diversity can be traced back at least to Aristotle. More recently, two successive revolutions rejuvenated this idea [1–3]: first, a proper mathematical refoundation of the theory of shape, then a technical revolution in the apparatus able to acquire raw data. By using a feature extraction method and planning its massive use on data acquired by aerial drones, the study by Lacaux and colleagues [4] retraces this curse of events.
The radial symmetry of Aurelia spp. jelly fish, a common species complex, is affected by stress and more largely by environmental variations, such as pollution exposition. Aurelia spp. normally present four gonads so that the proportion of non-tetramerous individuals in a population has been proposed as a biomarker [5,6].
In this study, the authors implemented the Hough transform to largely automate the detection of the gonads in Aurelia spp. Such use of the Hough transform, a long-used approach to identify shapes through edge detection, is new to morphometrics. Here, the Aurelia spp. gonads are identified as ellipses from which aspect descriptors can be derived, and primarily counted and thus can be used to quantify the proportion of individuals presenting body plans disorders.
The sample sizes studied here were too low to allow finer-grained ecophysiological investigations. That being said, the proof-of-concept is convincing and this paper paths the way for an operational and innovative approach to the ecological monitoring of sensible aquatic ecosystems.
References
[1] Kendall, D. G. (1989). A survey of the statistical theory of shape. Statistical Science, 87-99. doi: https://doi.org/10.1214/ss/1177012589
[2] Rohlf, F. J., and Marcus, L. F. (1993). A revolution morphometrics. Trends in ecology & evolution, 8(4), 129-132. doi: https://doi.org/10.1016/0169-5347(93)90024-J
[3] Adams, D. C., Rohlf, F. J., and Slice, D. E. (2004). Geometric morphometrics: ten years of progress following the ‘revolution’. Italian Journal of Zoology, 71(1), 5-16. doi: https://doi.org/10.1080/11250000409356545
[4] Lacaux, C., Desolneux, A., Gadreaud, J., Martin-Garin, B. and Thiéry, A. (2020) Hough transform implementation to evaluate the morphological variability of the moon jellyfish (Aurelia spp.). bioRxiv, 2020.03.11.986984, ver. 3 peer-reviewed and recommended by Peer Community in Ecology. doi: https://doi.org/10.1101/2020.03.11.986984
[5] Gershwin, L. A. (1999). Clonal and population variation in jellyfish symmetry. Journal of the Marine Biological Association of the United Kingdom, 79(6), 993-1000. doi: https://doi.org/10.1017/S0025315499001228
[6] Gadreaud, J., Martin-Garin, B., Artells, E., Levard, C., Auffan, M., Barkate, A.-L. and Thiéry, A. (2017) The moon jellyfish as a new bioindicator: impact of silver nanoparticles on the morphogenesis. In: Mariottini GL, editor. Jellyfish: ecology, distribution patterns and human interactions. Nova Science Publishers; 2017. pp. 277–292.
| Hough transform implementation to evaluate the morphological variability of the moon jellyfish (Aurelia spp.) | Céline Lacaux, Agnès Desolneux, Justine Gadreaud, Bertrand Martin-Garin and Alain Thiéry | <p>Variations of the animal body plan morphology and morphometry can be used as prognostic tools of their habitat quality. The potential of the moon jellyfish (Aurelia spp.) as a new model organism has been poorly tested. However, as a tetramerous... | 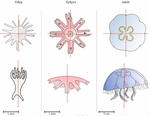 | Morphometrics | Vincent Bonhomme | | 2020-03-18 17:40:51 | View |
“Hidden” natural selection and the evolution of body size in harvested stocks
Recommended by Simon Blanchet based on reviews by Jean-François Arnoldi and 1 anonymous reviewer
Humans are exploiting biological resources since thousands of years. Exploitation of biological resources has become particularly intense since the beginning of the 20th century and the steep increase in the worldwide human population size. Marine and freshwater fishes are not exception to that rule, and they have been (and continue to be) strongly harvested as a source of proteins for humans. For some species, fishery has been so intense that natural stocks have virtually collapsed in only a few decades. The worst example begin that of the Northwest Atlantic cod that has declined by more than 95% of its historical biomasses in only 20-30 years of intensive exploitation (Frank et al. 2005). These rapid and steep changes in biomasses have huge impacts on the entire ecosystems since species targeted by fisheries are often at the top of trophic chains (Frank et al. 2005).
Beyond demographic impacts, fisheries also have evolutionary impacts on populations, which can also indirectly alter ecosystems (Uusi-Heikkilä et al. 2015; Palkovacs et al. 2018). Fishermen generally focus on the largest specimens, and hence exert a strong selective pressure against these largest fish (which is called “harvest selection”). There is now ample evidence that harvest selection can lead to rapid evolutionary changes in natural populations toward small individuals (Kuparinen & Festa-Bianchet 2017). These evolutionary changes are of course undesirable from a human perspective, and have attracted many scientific questions. Nonetheless, the consequence of harvest selection is not always observable in natural populations, and there are cases in which no phenotypic change (or on the contrary an increase in mean body size) has been observed after intense harvest pressures. In a conceptual Essay, Edeline and Loeuille (Edeline & Loeuille 2020) propose novel ideas to explain why the evolutionary consequences of harvest selection can be so diverse, and how a cross talk between ecological and evolutionary dynamics can explain patterns observed in natural stocks.
The general and novel concept proposed by Edeline and Loeuille is actually as old as Darwin’s book; The Origin of Species (Darwin 1859). It is based on the simple idea that natural selection acting on harvested populations can actually be strong, and counter-balance (or on the contrary reinforce) the evolutionary consequence of harvest selection. Although simple, the idea that natural and harvest selection are jointly shaping contemporary evolution of exploited populations lead to various and sometimes complex scenarios that can (i) explain unresolved empirical patterns and (ii) refine predictions regarding the long-term viability of exploited populations.
The Edeline and Loeuille’s crafty inspiration is that natural selection acting on exploited populations is itself an indirect consequence of harvest (Edeline & Loeuille 2020). They suggest that, by modifying the size structure of populations (a key parameter for ecological interactions), harvest indirectly alters interactions between populations and their biotic environment through competition and predation, which changes the ecological theatre and hence the selective pressures acting back to populations. They named this process “size-dependent eco-evolutionary feedback loops” and develop several scenarios in which these feedback loops ultimately deviate the evolutionary outcome of harvest selection from expectation. The scenarios they explore are based on strong theoretical knowledge, and range from simple ones in which a single species (the harvest species) is evolving to more complex (and realistic) ones in which multiple (e.g. the harvest species and its prey) species are co-evolving.
I will not come into the details of each scenario here, and I will let the readers (re-)discovering the complex beauty of biological life and natural selection. Nonetheless, I will emphasize the importance of considering these eco-evolutionary processes altogether to fully grasp the response of exploited populations. Edeline and Loeuille convincingly demonstrate that reduced body size due to harvest selection is obviously not the only response of exploited fish populations when natural selection is jointly considered (Edeline & Loeuille 2020). On the contrary, they show that –under some realistic ecological circumstances relaxing exploitative competition due to reduced population densities- natural selection can act antagonistically, and hence favour stable body size in exploited populations. Although this seems further desirable from a human perspective than a downsizing of exploited populations, it is actually mere window dressing as Edeline and Loeuille further showed that this response is accompanied by an erosion of the evolvability –and hence a lowest probability of long-term persistence- of these exploited populations.
Humans, by exploiting biological resources, are breaking the relative equilibrium of complex entities, and the response of populations to this disturbance is itself often complex and heterogeneous. In this Essay, Edeline and Loeuille provide –under simple terms- the theoretical and conceptual bases required to improve predictions regarding the evolutionary responses of natural populations to exploitation by humans (Edeline & Loeuille 2020). An important next step will be to generate data and methods allowing confronting the empirical reality to these novel concepts (e.g. (Monk et al. 2021), so as to identify the most likely evolutionary scenarios sustaining biological responses of exploited populations, and hence to set the best management plans for the long-term sustainability of these populations.
References
Darwin, C. (1859). On the Origin of Species by Means of Natural Selection. John Murray, London.
Edeline, E. & Loeuille, N. (2021) Size-dependent eco-evolutionary feedbacks in fisheries. bioRxiv, 2020.04.03.022905, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: https://doi.org/10.1101/2020.04.03.022905
Frank, K.T., Petrie, B., Choi, J. S. & Leggett, W.C. (2005). Trophic Cascades in a Formerly Cod-Dominated Ecosystem. Science, 308, 1621–1623. doi: https://doi.org/10.1126/science.1113075
Kuparinen, A. & Festa-Bianchet, M. (2017). Harvest-induced evolution: insights from aquatic and terrestrial systems. Philos. Trans. R. Soc. B Biol. Sci., 372, 20160036. doi: https://doi.org/10.1098/rstb.2016.0036
Monk, C.T., Bekkevold, D., Klefoth, T., Pagel, T., Palmer, M. & Arlinghaus, R. (2021). The battle between harvest and natural selection creates small and shy fish. Proc. Natl. Acad. Sci., 118, e2009451118. doi: https://doi.org/10.1073/pnas.2009451118
Palkovacs, E.P., Moritsch, M.M., Contolini, G.M. & Pelletier, F. (2018). Ecology of harvest-driven trait changes and implications for ecosystem management. Front. Ecol. Environ., 16, 20–28. doi: https://doi.org/10.1002/fee.1743
Uusi-Heikkilä, S., Whiteley, A.R., Kuparinen, A., Matsumura, S., Venturelli, P.A., Wolter, C., et al. (2015). The evolutionary legacy of size-selective harvesting extends from genes to populations. Evol. Appl., 8, 597–620. doi: https://doi.org/10.1111/eva.12268
| Size-dependent eco-evolutionary feedbacks in fisheries | Eric Edeline and Nicolas Loeuille | <p>Harvesting may drive body downsizing along with population declines and decreased harvesting yields. These changes are commonly construed as direct consequences of harvest selection, where small-bodied, early-reproducing individuals are immedia... | 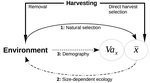 | Biodiversity, Community ecology, Competition, Eco-evolutionary dynamics, Evolutionary ecology, Food webs, Interaction networks, Life history, Population ecology, Theoretical ecology | Simon Blanchet | | 2020-04-03 16:14:05 | View |
Insect herbivory on urban trees: Complementary effects of tree neighbours and predation
Alex Stemmelen, Alain Paquette, Marie-Lise Benot, Yasmine Kadiri, Hervé Jactel, Bastien Castagneyrol
https://doi.org/10.1101/2020.04.15.042317
Tree diversity is associated with reduced herbivory in urban forest
Recommended by Ruth Arabelle Hufbauer and Ian Pearse based on reviews by Ian Pearse and Freerk Molleman
Urban ecology, the study of ecological systems in our increasingly urbanized world, is crucial to planning and redesigning cities to enhance ecosystem services (Kremer et al. 2016), human health and well-being and further conservation goals (Dallimer et al. 2012). Urban trees are a crucial component of urban streets and parks that provide shade and cooling through evapotranspiration (Fung and Jim 2019), improve air quality (Lai and Kontokosta 2019), help control storm water (Johnson and Handel 2016), and conserve wildlife (Herrmann et al. 2012; de Andrade et al. 2020).
Ideally, management of urban forests strikes a balance between maintaining the health of urban trees while retaining those organisms, such as herbivores, that connect a tree to the urban ecosystem. Herbivory by arthropods can substantially affect tree growth and reproduction (Whittaker and Warrington 1985), and so understanding factors that influence herbivory in urban forests is important to effective management. At the same time, herbivorous arthropods are important as key components of urban bird diets (Airola and Greco 2019) and provide a backyard glimpse at forest ecosystems in an increasingly built environment (Pearse 2019). Maintenance of arthropod predators may be one way to retain arthropods in urban forests while keeping detrimental outbreaks of herbivores in check. In “Insect herbivory on urban trees: Complementary effects of tree neighbors and predation” Stemmelen and colleagues (Stemmelen et al. 2020) use a clever sampling design to show that insect herbivory decreases as the diversity of neighboring trees increased. By placing artificial larvae out on trees, they provide evidence that increased predation in higher diversity urban forest patches might drive patterns in herbivory. The paper also demonstrates the importance of tree species identity in determining leaf herbivory.
The implications of this research for urban foresters is that deliberately planting diverse urban forests will help manage insect herbivores and should thus improve tree health. Potential knock-on effects could be seen for the ecosystem services provided by urban forests. While it might be tempting to simply plant more of the species that are subject to low current rates of herbivory, other research on the long-term vulnerability of monocultures to attack by specialist pathogens and herbivores (Tooker and Frank 2012) cautions against such an approach. Furthermore, the importance of urban forest insects to birds, including migrating birds, argues for managing urban forests more holistically (Greco and Airola 2018).
Stemmelen et al. (2020) used an observational approach focused on urban forests in Montreal, Canada in their research. Their findings suggest follow-up research focused on a broader cross-section of urban forests across latitudes, as well as experimental research. Experiments could, for example, exclude avian predators with netting (e.g. (Marquis and Whelan 1994)) to evaluate the relative importance of birds to managing urban insects on trees, as well as the flip side of that equation, the important to birds of insects on urban trees.
In summary, Stemmelen and colleague’s manuscript illustrates clever sampling and use of observational data to infer broader ecological patterns. It is worth reading to better understand the role of diversity in driving plant-insect community interactions and given the implications of the findings for sustainable long-term management of urban forests.
References
Airola, D. and Greco, S. (2019). Birds and oaks in California’s urban forest. Int. Oaks, 30, 109–116.
de Andrade, A.C., Medeiros, S. and Chiarello, A.G. (2020). City sloths and marmosets in Atlantic forest fragments with contrasting levels of anthropogenic disturbance. Mammal Res., 65, 481–491. doi: https://doi.org/10.1007/s13364-020-00492-0
Dallimer, M., Irvine, K.N., Skinner, A.M.J., Davies, Z.G., Rouquette, J.R., Maltby, L.L., et al. (2012). Biodiversity and the Feel-Good Factor: Understanding Associations between Self-Reported Human Well-being and Species Richness. Bioscience, 62, 47–55. doi: https://doi.org/10.1525/bio.2012.62.1.9
Fung, C.K.W. and Jim, C.Y. (2019). Microclimatic resilience of subtropical woodlands and urban-forest benefits. Urban For. Urban Green., 42, 100–112. doi: https://doi.org/10.1016/j.ufug.2019.05.014
Greco, S.E. and Airola, D.A. (2018). The importance of native valley oaks (Quercus lobata) as stopover habitat for migratory songbirds in urban Sacramento, California, USA. Urban For. Urban Green., 29, 303–311. doi: https://doi.org/10.1016/j.ufug.2018.01.005
Herrmann, D.L., Pearse, I.S. and Baty, J.H. (2012). Drivers of specialist herbivore diversity across 10 cities. Landsc. Urban Plan., 108, 123–130. doi: https://doi.org/10.1016/j.landurbplan.2012.08.007
Johnson, L.R. and Handel, S.N. (2016). Restoration treatments in urban park forests drive long-term changes in vegetation trajectories. Ecol. Appl., 26, 940–956. doi: https://doi.org/10.1890/14-2063
Kremer, P., Hamstead, Z., Haase, D., McPhearson, T., Frantzeskaki, N., Andersson, E., et al. (2016). Key insights for the future of urban ecosystem services research. Ecol. Soc., 21: 29. doi: http://doi.org/10.5751/ES-08445-210229
Lai, Y. and Kontokosta, C.E. (2019). The impact of urban street tree species on air quality and respiratory illness: A spatial analysis of large-scale, high-resolution urban data. Heal. Place, 56, 80–87. doi: https://doi.org/10.1016/j.healthplace.2019.01.016
Marquis, R.J. and Whelan, C.J. (1994). Insectivorous birds increase growth of white oak through consumption of leaf-chewing insects. Ecology, 75, 2007–2014. doi: https://doi.org/10.2307/1941605
Pearse, I.S. (2019). Insect herbivores on urban native oak trees. Int. Oaks, 30, 101–108.
Stemmelen, A., Paquette, A., Benot, M.-L., Kadiri, Y., Jactel, H. and Castagneyrol, B. (2020) Insect herbivory on urban trees: Complementary effects of tree neighbours and predation. bioRxiv, 2020.04.15.042317, ver. 5 peer-reviewed and recommended by PCI Ecology. doi: https://doi.org/10.1101/2020.04.15.042317
Tooker, J. F., and Frank, S. D. (2012). Genotypically diverse cultivar mixtures for insect pest management and increased crop yields. J. Appl. Ecol., 49(5), 974-985. doi: https://doi.org/10.1111/j.1365-2664.2012.02173.x
Whittaker, J.B. and Warrington, S. (1985). An experimental field study of different levels of insect herbivory induced By Formica rufa predation on Sycamore (Acer pseudoplatanus) III. Effects on Tree Growth. J. Appl. Ecol., 22, 797. doi: https://doi.org/10.2307/2403230
| Insect herbivory on urban trees: Complementary effects of tree neighbours and predation | Alex Stemmelen, Alain Paquette, Marie-Lise Benot, Yasmine Kadiri, Hervé Jactel, Bastien Castagneyrol | <p>Insect herbivory is an important component of forest ecosystems functioning and can affect tree growth and survival. Tree diversity is known to influence insect herbivory in natural forest, with most studies reporting a decrease in herbivory wi... |  | Biodiversity, Biological control, Community ecology, Ecosystem functioning, Herbivory | Ruth Arabelle Hufbauer | | 2020-04-20 13:49:36 | View |
How citizen science could improve Species Distribution Models and their independent assessment
Florence Matutini, Jacques Baudry, Guillaume Pain, Morgane Sineau, Josephine Pithon
https://doi.org/10.1101/2020.06.02.129536
Citizen science contributes to SDM validation
Recommended by Francisco Lloret based on reviews by Maria Angeles Perez-Navarro and 1 anonymous reviewer
Citizen science is becoming an important piece for the acquisition of scientific knowledge in the fields of natural sciences, and particularly in the inventory and monitoring of biodiversity (McKinley et al. 2017). The information generated with the collaboration of citizens has an evident importance in conservation, by providing information on the state of populations and habitats, helping in mitigation and restoration actions, and very importantly contributing to involve society in conservation (Brown and Williams 2019).
An obvious advantage of these initiatives is the ability to mobilize human resources on a large territorial scale and in the medium term, which would otherwise be difficult to finance. The resulting increasing information then can be processed with advanced computational techniques (Hochachka et al 2012; Kelling et al. 2015), thus improving our interpretation of the distribution of species. Specifically, the ability to obtain information on a large territorial scale can be integrated into studies based on Species Distribution Models SDMs. One of the common problems with SDMs is that they often work from species occurrences that have been opportunistically recorded, either by professionals or amateurs. A great challenge for data obtained from non-professional citizens, however, remains to ensure its standardization and quality (Kosmala et al. 2016). This requires a clear and effective design, solid volunteer training, and a high level of coordination that turns out to be complex (Brown and Williams 2019). Finally, it is essential to perform a quality validation following scientifically recognized standards, since they are often conditioned by errors and biases in obtaining information (Bird et al. 2014). There are two basic approaches to obtain the necessary data for this validation: getting it from an external source (external validation), or allocating a part of the database itself (internal validation or cross-validation) to this function.
Matutini et al. (2020) in his work 'How citizen science could improve Species Distribution Models and their independent assessment' shows a novel application of the data generated by a citizen science initiative ('Un Dragon dans mon Jardin') by providing an external source for the validation of SDMs, as a tool to construct habitat suitability maps for nine species of amphibians in western France. Importantly, 'Un Dragon dans mon Jardin' contains standardized presence-absence data, the approximation recognized as the most robust (Guisan, et al. 2017). The SDMs to be validated, in turn, were based on opportunistic information obtained by citizens and professionals. The result shows the usefulness of this external data source by minimizing the overestimation of model accuracy that is obtained with cross-validation with the internal evaluation dataset. It also shows the importance of properly filtering the information obtained by citizens by determining the threshold of sampling effort.
The destiny of citizen science is to be integrated into the complex world of science. Supported by the increasing level of the formation of society, it is becoming a fundamental piece in the scientific system dedicated to the study of biodiversity and its conservation. After funding for scientists specialized in the recognition of biodiversity has been cut back, we are seeing a transformation of the activity of these scientists towards the design, coordination, training and verification of programs for the acquisition of field information obtained by citizens. A main goal is that a substantial part of this information will eventually get integrated into the scientific system, and rigorous verification process a fundamental element for such purpose, as shown by Matutini et al. (2020) work.
References
[1] Bird TJ et al. (2014) Statistical solutions for error and bias in global citizen science datasets. Biological Conservation 173: 144-154. doi: 10.1016/j.biocon.2013.07.037
[2] Brown ED and Williams BK (2019) The potential for citizen science to produce reliable and useful information in ecology. Conservation Biology 33: 561-569. doi: 10.1111/cobi.13223
[3] Guisan A, Thuiller W and Zimmermann N E (2017) Habitat Suitability and Distribution Models: With Applications in R. The University of Chicago Press. doi: 10.1017/9781139028271
[4] Hochachka WM, Fink D, Hutchinson RA, Sheldon D, Wong WK and Kelling S (2012) Data-intensive science applied to broad-scale citizen science. Trens Ecol Evol 27: 130-137. doi: 10.1016/j.tree.2011.11.006
[5] Kelling S, Fink D, La Sorte FA, Johnston A, Bruns NE and Hochachka WM (2015) Taking a ‘Big Data’ approach to data quality in a citizen science project. Ambio 44(Supple. 4):S601-S611. doi: 10.1007/s13280-015-0710-4
[6] Kosmala M, Wiggins A, Swanson A and Simmons B (2016) Assessing data quality in citizen science. Front Ecol Environ 14: 551–560. doi: 10.1002/fee.1436
[7] Matutini F, Baudry J, Pain G, Sineau M and Pithon J (2020) How citizen science could improve Species Distribution Models and their independent assessment. bioRxiv, 2020.06.02.129536, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/2020.06.02.129536
[8] McKinley DC et al. (2017) Citizen science can improve conservation science, natural resource management, and environmental protection. Biological Conservation 208:15-28. doi: 10.1016/j.biocon.2016.05.015
| How citizen science could improve Species Distribution Models and their independent assessment | Florence Matutini, Jacques Baudry, Guillaume Pain, Morgane Sineau, Josephine Pithon | <p>Species distribution models (SDM) have been increasingly developed in recent years but their validity is questioned. Their assessment can be improved by the use of independent data but this can be difficult to obtain and prohibitive to collect.... |  | Biodiversity, Biogeography, Conservation biology, Habitat selection, Spatial ecology, Metacommunities & Metapopulations, Species distributions, Statistical ecology | Francisco Lloret | | 2020-06-03 09:36:34 | View |
The dynamics of spawning acts by a semelparous fish and its associated energetic costs
Cédric Tentelier, Colin Bouchard, Anaïs Bernardin, Amandine Tauzin, Jean-Christophe Aymes, Frédéric Lange, Charlotte Recapet, Jacques Rives
https://doi.org/10.1101/436295
Extreme weight loss: when accelerometer could reveal reproductive investment in a semelparous fish species
Recommended by Francois-Xavier Dechaume-Moncharmont based on reviews by Aidan Jonathan Mark Hewison, Loïc Teulier and 1 anonymous reviewer
Continuous observation of animal behaviour could be quite a challenge in the field, and the situation becomes even more complicated with aquatic species mostly active at night. In such cases, biologging techniques are real game changers in ecology, behavioural ecology or eco-physiology. An accelerating number of methodological applications of these tools in natural condition are thus published each year [1]. Biologging is not limited to movement ecology. For instance, fine grain information about energy expenditure can be inferred from body acceleration [2], and accelerometers has already proven useful in monitoring reproductive costs in some fish species [3,4]. The first part of the study by Tentelier et al. [5] is in line with this growing literature. It describes measurements of energy expenditure during reproduction in a fish species, Allis shad (Alosa Alosa), based on tail beat frequency and occurrence of spawning acts. The study has been convincingly conducted, and the results are important for fish biologists. But this is not the whole story: the authors added to this otherwise classical study a very original and insightful analysis which deserves closer interest.
Tentelier et al. propose to use static accelerometer to monitor change in body roundness through the reproductive season. These semelparous fish first mature and built up reserves in the Atlantic Ocean and migrate into fresh water to reproduce. Contrary to iteroparous species, female shads do not have to strategically preserve energy for future reproduction. The females die few days after spawning having exhausted their energetic reserves: they typically lose almost half of their body mass during the spawning season. The beautiful idea in this study was to track down information about this dramatic slimming in the accelerometer data. Indeed, the accelerometer was attached on the side of the fish (close to the dorsal fin). A change in its angle with the vertical plane could be correlated with the change in roundness, the angle declining with the female thinning. Accelerometers have already been used to record body posture [6] but, in the present study, the novelty was to monitor the change in body shape.
Unfortunately, the data by Tentelier et al. are inconclusive so far. Broadly speaking, the accelerometer angle recorded declined through the spawning season, indicating an average slimming of the females, but there was no correlation between the change in angle and the mass loss at the individual level. This was partly due to the fact that the dorsal position of the accelerometer was not optimized to measures egg laying whose effects are mostly observable on ventral side.
Yet, this nice idea deserves more scrutiny. The method seems to be sensitive enough to detect inflation of swim bladder, the gas-filled organ helping the fish to control their position in the water column, as the accelerometer angle increased when the fish stayed close to the water surface. Additional works and proper calibration are certainly needed to validate the use of accelerometer angle as a proxy for body roundness. The actual data were not strong enough to justify a standalone publication on the subject, but it would have been shame to lose traces of such analysis and keep it in the file drawer. This is why I strongly support its report as a side question in a broader study. Science progresses not only with neat conclusive studies but also when unexpected (apparently anecdotal) observations stimulate new researches.
References
[1] Börger L, Bijleveld AI, Fayet AL, Machovsky‐Capuska GE, Patrick SC, Street GM and Vander Wal E. (2020) Biologging special feature. J. Anim. Ecol. 89, 6–15. 10.1111/1365-2656.13163
[2] Wilson RP et al. (2020) Estimates for energy expenditure in free‐living animals using acceleration proxies: A reappraisal. J. Anim. Ecol. 89, 161–172. 10.1111/1365-2656.13040
[3] Tsuda Y, Kawabe R, Tanaka H, Mitsunaga Y, Hiraishi T, Yamamoto K and Nashimoto K. (2006) Monitoring the spawning behaviour of chum salmon with an acceleration data logger. Ecol. Freshw. Fish 15, 264–274. 10.1111/j.1600-0633.2006.00147.x
[4] Sakaji H, Hamada K, Naito Y. 2018 Identifying spawning events of greater amberjack using accelerometers. Mar. Biol. Res. 14, 637–641. 10.1080/17451000.2018.1492140
[5] Tentelier C, Bouchard C, Bernardin A, Tauzin A, Aymes J-C, Lange F, Récapet C, Rives J (2020) The dynamics of spawning acts by a semelparous fish and its associated energetic costs. bioRxiv, 436295. doi: 10.1101/436295 ver. 7 peer-reviewed and recommended by PCI Ecology. 10.1101/436295
[6] Brown DD, Kays R, Wikelski M, Wilson R, Klimley AP. 2013 Observing the unwatchable through acceleration logging of animal behavior. Anim. Biotelemetry 1, 20. 10.1186/2050-3385-1-20
| The dynamics of spawning acts by a semelparous fish and its associated energetic costs | Cédric Tentelier, Colin Bouchard, Anaïs Bernardin, Amandine Tauzin, Jean-Christophe Aymes, Frédéric Lange, Charlotte Recapet, Jacques Rives | <p>1. During the reproductive season, animals have to manage both their energetic budget and gamete stock. In particular, for semelparous capital breeders with determinate fecundity and no parental care other than gametic investment, the depletion... |  | Behaviour & Ethology, Freshwater ecology, Life history | Francois-Xavier Dechaume-Moncharmont | | 2020-06-04 15:18:56 | View |
Assessing bat-vehicle collision risks using acoustic 3D tracking
Recommended by Gloriana Chaverri based on reviews by Mark Brigham and ?
The loss of biodiversity is an issue of great concern, especially if the extinction of species or the loss of a large number of individuals within populations results in a loss of critical ecosystem services. We know that the most important threat to most species is habitat loss and degradation (Keil et al., 2015; Pimm et al., 2014); the latter can be caused by multiple anthropogenic activities, including pollution, introduction of invasive species and fragmentation (Brook et al., 2008; Scanes, 2018). Roads are a major cause of habitat fragmentation, isolating previously connected populations and being a direct source of mortality for animals that attempt to cross them (Spellberg, 1998).
While most studies have focused on the effect of roads on larger mammals (Bartonička et al., 2018; Litvaitis and Tash, 2008), in recent years many researchers have grown increasingly concerned about the risk of collision between bats and vehicles (Fensome and Mathews, 2016). For example, a recent publication by Medinas et al. (2021) found 509 bat casualties along a 51-km-long transect during a period of 3 years. Their study provides extremely valuable information to asses which factors primarily drive bat mortality on roads, yet it required a substantial investment of time coupled with the difficulty of detecting bat carcasses. Other studies have used acoustic monitoring as a proxy to gauge risk of collision based on estimates of bat density along roads (reviewed in Fensome and Mathews 2016); while the results of such studies are valuable, the number of passes recorded does not necessarily equal collision risk, as many species may simply avoid crossing the roads. Understanding the risk of collisions is of vital importance for adequate planning of road construction, particularly for key sites that harbor threatened bat species or unusually large populations, especially if these are already greatly impacted by other anthropogenic activities (e.g. wind turbines; Kunz et al. 2007) or unusually deadly pathogens (e.g. white-nose syndrome; Blehert et al. 2009).
The study by Roemer et al. (2020) titled “Influence of local landscape and time of year on bat-road collision risks”, is a welcome addition to our understanding of bat collision risk as it employs a more accurate assessment of bat collision risk based on acoustic monitoring and tracking of flight paths. The goal of the study of Roemer and collaborators, which was conducted at 66 study sites in the Mediterranean region, is to provide an assessment of collision risk based on bat activity near roads. They collected a substantial amount of information for several species: more than 30,000 estimated flight trajectories for 21+ species, including Barbastella barbastellus, Myotis spp., Plecotus sp., Rhinolophus ferrumequinum, Miniopterus schreibersii, Pipistrellus spp., Nyctalus leisleri, and others. They assess risk based on estimates of 1) species abundance from acoustic monitoring, 2) direction of flight paths along roads, and 3) bat-vehicle co-occurrence.
Their findings suggest that risk is habitat, species, guild, and season-specific. Roads within forested habitats posed the largest threats for most species, particularly since most flights within these habitats occurred at the zone of collision risk. They also found that bats typically fly parallel to the road axis regardless of habitat type, which they argue supports the idea that bats may use roads as corridors. The results of their study, as expected, also show that the majority of bat passes were detected during summer or autumn, depending on species, yet they provide novel findings of an increase in risky behaviors during autumn, when the number of passes at the zone of collision risk increased significantly. Their results also suggest that mid-range echolocators, a classification that is based on call design and parameters (Frey-Ehrenbold et al., 2013), had a larger portion of flights in the zone at risk, thus potentially making them more susceptible than short and long-range echolocators to collisions with vehicles.
The methods employed by Roemer et al. (2020) could further help us determine how roads pose species and site-specific threats in a diversity of places without the need to invest a significant amount of time locating bat carcasses. Their findings are also important as they could provide valuable information for deciding where new roads should be constructed, particularly if the most vulnerable species are abundant, perhaps due to the presence of important roost sites. They also show how habitats near larger roads could increase threats, providing an important first step for recommendations regarding road construction and maintenance. As pointed out by one reviewer, one possible limitation of the study is that the results are not supported by the identification of carcasses. For example, does an increase in the number of identified flights at the zone of risk really translate into an increase in the number of collisions? Regardless of the latter, the paper’s methods and results are very valuable and provide an important step towards developing additional tools to assess bat-vehicle collision risks.
References
[1] Bartonička T, Andrášik R, Duľa M, Sedoník J, Bíl M (2018) Identification of local factors causing clustering of animal-vehicle collisions. The Journal of Wildlife Management, 82, 940–947. https://doi.org/10.1002/jwmg.21467
[2] Blehert DS, Hicks AC, Behr M, Meteyer CU, Berlowski-Zier BM, Buckles EL, Coleman JTH, Darling SR, Gargas A, Niver R, Okoniewski JC, Rudd RJ, Stone WB (2009) Bat White-Nose Syndrome: An Emerging Fungal Pathogen? Science, 323, 227–227. https://doi.org/10.1126/science.1163874
[3] Brook BW, Sodhi NS, Bradshaw CJA (2008) Synergies among extinction drivers under global change. Trends in Ecology & Evolution, 23, 453–460. https://doi.org/10.1016/j.tree.2008.03.011
[4] Fensome AG, Mathews F (2016) Roads and bats: a meta-analysis and review of the evidence on vehicle collisions and barrier effects. Mammal Review, 46, 311–323. https://doi.org/10.1111/mam.12072
[5] Frey‐Ehrenbold A, Bontadina F, Arlettaz R, Obrist MK (2013) Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matrices. Journal of Applied Ecology, 50, 252–261. https://doi.org/10.1111/1365-2664.12034
[6] Keil P, Storch D, Jetz W (2015) On the decline of biodiversity due to area loss. Nature Communications, 6, 8837. https://doi.org/10.1038/ncomms9837
[7] Kunz TH, Arnett EB, Erickson WP, Hoar AR, Johnson GD, Larkin RP, Strickland MD, Thresher RW, Tuttle MD (2007) Ecological impacts of wind energy development on bats: questions, research needs, and hypotheses. Frontiers in Ecology and the Environment, 5, 315–324. https://doi.org/10.1890/1540-9295(2007)5[315:EIOWED]2.0.CO;2
[8] Litvaitis JA, Tash JP (2008) An Approach Toward Understanding Wildlife-Vehicle Collisions. Environmental Management, 42, 688–697. https://doi.org/10.1007/s00267-008-9108-4
[9] Medinas D, Marques JT, Costa P, Santos S, Rebelo H, Barbosa AM, Mira A (2021) Spatiotemporal persistence of bat roadkill hotspots in response to dynamics of habitat suitability and activity patterns. Journal of Environmental Management, 277, 111412. https://doi.org/10.1016/j.jenvman.2020.111412
[10] Pimm SL, Jenkins CN, Abell R, Brooks TM, Gittleman JL, Joppa LN, Raven PH, Roberts CM, Sexton JO (2014) The biodiversity of species and their rates of extinction, distribution, and protection. Science, 344. https://doi.org/10.1126/science.1246752
[11] Roemer C, Coulon A, Disca T, Bas Y (2020) Influence of local landscape and time of year on bat-road collision risks. bioRxiv, 2020.07.15.204115, ver. 3 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2020.07.15.204115
[12] Scanes CG (2018) Chapter 19 - Human Activity and Habitat Loss: Destruction, Fragmentation, and Degradation. In: Animals and Human Society (eds Scanes CG, Toukhsati SR), pp. 451–482. Academic Press. https://doi.org/10.1016/B978-0-12-805247-1.00026-5
[13] Spellerberg I (1998) Ecological effects of roads and traffic: a literature review. Global Ecology & Biogeography Letters, 7, 317–333. https://doi.org/10.1046/j.1466-822x.1998.00308.x
| Influence of local landscape and time of year on bat-road collision risks | Charlotte Roemer, Aurélie Coulon, Thierry Disca, and Yves Bas | <p>Roads impact bat populations through habitat loss and collisions. High quality habitats particularly increase bat mortalities on roads, yet many questions remain concerning how local landscape features may influence bat behaviour and lead to hi... |  | Behaviour & Ethology, Biodiversity, Conservation biology, Human impact, Landscape ecology | Gloriana Chaverri | | 2020-07-20 10:56:29 | View |
Once upon a time in the far south: Influence of local drivers and functional traits on plant invasion in the harsh sub-Antarctic islands
Manuele Bazzichetto, François Massol, Marta Carboni, Jonathan Lenoir, Jonas Johan Lembrechts, Rémi Joly, David Renault
https://doi.org/10.1101/2020.07.19.210880
A meaningful application of species distribution models and functional traits to understand invasion dynamics
Recommended by Joaquín Hortal based on reviews by Paula Matos and Peter Convey
Polar and subpolar regions are fragile environments, where the introduction of alien species may completely change ecosystem dynamics if the alien species become keystone species (e.g. Croll, 2005). The increasing number of human visits, together with climate change, are favouring the introduction and settling of new invaders to these regions, particularly in Antarctica (Hughes et al. 2015). Within this context, the joint use of Species Distribution Models (SDM) –to assess the areas potentially suitable for the aliens– with other measures of the potential to become successful invaders can inform on the need for devoting specific efforts to eradicate these new species before they become naturalized (e.g. Pertierra et al. 2016).
Bazzichetto et al. (2020) use data from a detailed inventory, SDMs and trait data altogether to assess the drivers of invasion success of six alien plants on Possession Island, in the remote sub-Antarctic archipelago of Crozet. SDMs have inherent limitations to describe different aspects of species distributions, including the fundamental niche and, with it, the areas that could host viable populations (Hortal et al. 2012). Therefore, their utility to predict future biological invasions is limited (Jiménez-Valverde et al. 2011). However, they can be powerful tools to describe species range dynamics if they are thoughtfully used by adopting conscious decisions about the techniques and data used, and interpreting carefully the actual implications of their results.
This is what Bazzichetto et al. (2020) do, using General Linear Models (GLM) –a technique well rooted in the original niche-based SDM theory (e.g. Austin 1990)– that can provide a meaningful description of the realized niche within the limits of an adequately sampled region. Further, as alien species share and are similarly affected by several steps of the invasion process (Richardson et al. 2000), these authors model the realized distribution of the six species altogether. This can be done through the recently developed joint-SDM, a group of techniques where the co-occurrence of the modelled species is explicitly taken into account during modelling (e.g. Pollock et al. 2014). Here, the addition of species traits has been identified as a key step to understand the associations of species in space (see Dormann et al. 2018). Bazzichetto et al. (2020) combine their GLM-based SDM for each species with a so-called multi-SDM approach, where they assess together the consistency in the interactions between both species and topographically-driven climate variations, and several plant traits and two key anthropic factors –accessibility from human settlements and distance to hiking paths.
This work is a good example on how a theoretically meaningful SDM approach can provide useful –though perhaps not deep– insights on biological invasions for remote landscapes threatened by biotic homogenization. By combining climate and topographic variables as proxies for the spatial variations in the abiotic conditions regulating plant growth, measures of accessibility, and traits of the plant invaders, Bazzichetto et al. (2020) are able to identify the different effects that the interactions between the potential intensity of propagule dissemination by humans, and the ecological characteristics of the invaders themselves, may have on their invasion success.
The innovation of modelling together species responses is important because it allows dissecting the spatial dynamics of spread of the invaders, which indeed vary according to a handful of their traits. For example, their results show that no all old residents have profited from the larger time of residence in the island, as Poa pratensis is seemingly as dependent of a higher intensity of human activity as the newcomer invaders in general are. According to Bazzichetto et al. trait-based analyses, these differences are apparently related with plant height, as smaller plants disperse more easily. Further, being perennial also provides an advantage for the persistence in areas with less human influence. This puts name, shame and fame to the known influence of plant life history on their dispersal success (Beckman et al. 2018), at least for the particular case of plant invasions in Possession Island.
Of course this approach has limitations, as data on the texture, chemistry and temperature of the soil are not available, and thus were not considered in the analyses. These factors may be critical for both establishment and persistence of small plants in the harsh Antarctic environments, as Bazzichetto et al. (2020) recognize. But all in all, their results provide key insights on which traits may confer alien plants with a higher likelihood of becoming successful invaders in the fragile Antarctic and sub-Antarctic ecosystems. This opens a way for rapid assessments of invasibility, which will help identifying which species in the process of naturalizing may require active contention measures to prevent them from becoming ecological game changers and cause disastrous cascade effects that shift the dynamics of native ecosystems.
References
Austin, M. P., Nicholls, A. O., and Margules, C. R. (1990). Measurement of the realized qualitative niche: environmental niches of five Eucalyptus species. Ecological Monographs, 60(2), 161-177. doi: https://doi.org/10.2307/1943043
Bazzichetto, M., Massol, F., Carboni, M., Lenoir, J., Lembrechts, J. J. and Joly, R. (2020) Once upon a time in the far south: Influence of local drivers and functional traits on plant invasion in the harsh sub-Antarctic islands. bioRxiv, 2020.07.19.210880, ver. 3 peer-reviewed and recommended by PCI Ecology. doi: https://doi.org/10.1101/2020.07.19.210880
Beckman, N. G., Bullock, J. M., and Salguero-Gómez, R. (2018). High dispersal ability is related to fast life-history strategies. Journal of Ecology, 106(4), 1349-1362. doi: https://doi.org/10.1111/1365-2745.12989
Croll, D. A., Maron, J. L., Estes, J. A., Danner, E. M., and Byrd, G. V. (2005). Introduced predators transform subarctic islands from grassland to tundra. Science, 307(5717), 1959-1961. doi: https://doi.org/10.1126/science.1108485
Dormann, C. F., Bobrowski, M., Dehling, D. M., Harris, D. J., Hartig, F., Lischke, H., Moretti, M. D., Pagel, J., Pinkert, S., Schleuning, M., Schmidt, S. I., Sheppard, C. S., Steinbauer, M. J., Zeuss, D., and Kraan, C. (2018). Biotic interactions in species distribution modelling: 10 questions to guide interpretation and avoid false conclusions. Global Ecology and Biogeography, 27(9), 1004-1016. doi: https://doi.org/10.1111/geb.12759
Jiménez-Valverde, A., Peterson, A., Soberón, J., Overton, J., Aragón, P., and Lobo, J. (2011). Use of niche models in invasive species risk assessments. Biological Invasions, 13(12), 2785-2797. doi: https://doi.org/10.1007/s10530-011-9963-4
Hortal, J., Lobo, J. M., and Jiménez-Valverde, A. (2012). Basic questions in biogeography and the (lack of) simplicity of species distributions: Putting species distribution models in the right place. Natureza & Conservação – Brazilian Journal of Nature Conservation, 10(2), 108-118. doi: https://doi.org/10.4322/natcon.2012.029
Hughes, K. A., Pertierra, L. R., Molina-Montenegro, M. A., and Convey, P. (2015). Biological invasions in terrestrial Antarctica: what is the current status and can we respond? Biodiversity and Conservation, 24(5), 1031-1055. doi: https://doi.org/10.1007/s10531-015-0896-6
Pertierra, L. R., Baker, M., Howard, C., Vega, G. C., Olalla-Tarraga, M. A., and Scott, J. (2016). Assessing the invasive risk of two non-native Agrostis species on sub-Antarctic Macquarie Island. Polar Biology, 39(12), 2361-2371. doi: https://doi.org/10.1007/s00300-016-1912-3
Pollock, L. J., Tingley, R., Morris, W. K., Golding, N., O'Hara, R. B., Parris, K. M., Vesk, P. A., and McCarthy, M. A. (2014). Understanding co-occurrence by modelling species simultaneously with a Joint Species Distribution Model (JSDM). Methods in Ecology and Evolution, 5(5), 397-406. doi: https://doi.org/10.1111/2041-210X.12180
Richardson, D. M., Pyšek, P., Rejmánek, M., Barbour, M. G., Panetta, F. D., and West, C. J. (2000). Naturalization and invasion of alien plants: concepts and definitions. Diversity and Distributions, 6(2), 93-107. doi: https://doi.org/10.1046/j.1472-4642.2000.00083.x
| Once upon a time in the far south: Influence of local drivers and functional traits on plant invasion in the harsh sub-Antarctic islands | Manuele Bazzichetto, François Massol, Marta Carboni, Jonathan Lenoir, Jonas Johan Lembrechts, Rémi Joly, David Renault | <p>Aim Here, we aim to: (i) investigate the local effect of environmental and human-related factors on alien plant invasion in sub-Antarctic islands; (ii) explore the relationship between alien species features and their dependence on anthropogeni... |  | Biogeography, Biological invasions, Spatial ecology, Metacommunities & Metapopulations, Species distributions | Joaquín Hortal | | 2020-07-21 21:13:08 | View |







