Trophic niche of the invasive gregarious species Crepidula fornicata, in relation to ontogenic changes
Thibault Androuin, Stanislas F. Dubois, Cédric Hubas, Gwendoline Lefebvre, Fabienne Le Grand, Gauthier Schaal, Antoine Carlier
https://doi.org/10.1101/2020.07.30.229021
A lack of clear dietary differences between ontogenetic stages of invasive slippersnails provides important insights into resource use and potential inter- and intra-specific competition
Recommended by Matthew Bracken based on reviews by 2 anonymous reviewers
The slippersnail (Crepidula fornicata), originally from the eastern coast of North America, has invaded European coastlines from Norway to the Mediterranean Sea [1]. This species is capable of achieving incredibly high densities (up to several thousand individuals per square meter) and likely has major impacts on a variety of community- and ecosystem-level processes, including alteration of carbon and nitrogen fluxes and competition with native suspension feeders [2].
Given this potential for competition, it is important to understand the diet of C. fornicata and its potential overlap with native species. However, previous research on the diet of C. fornicata and related species suggests that the types of food consumed may change with age [3, 4]. This species has an unusual reproductive strategy. It is a sequential hermaphrodite, which begins life as a somewhat mobile male but eventually slows down to become sessile. Sessile individuals form stacks of up to 10 or more individuals, with larger individuals on the bottom of the stack, and decreasingly smaller individuals piled on top. Snails at the bottom of the stack are female, whereas snails at the top of the stack are male; when the females die, the largest males become female [5]. Thus, understanding these potential ontogenetic dietary shifts has implications for both intraspecific (juvenile vs. male vs. female) and interspecific competition associated with an abundant, invasive species.
To this end, Androuin and colleagues evaluated the stable-isotope (d13C and d15N) and fatty-acid profiles of food sources and different life-history stages of C. fornicata [6]. Based on previous work highlighting the potential for life-history changes in the diet of this species [3,4], they hypothesized that C. fornicata would shift its diet as it aged and predicted that this shift would be reflected in changes in its stable-isotope and fatty-acid profiles. The authors found that potential food sources (biofilm, suspended particulate organic matter, and superficial sedimentary organic matter) differed substantially in both stable-isotope and fatty-acid signatures. However, whereas fatty-acid profiles changed substantially with age, there was no shift in the stable-isotope signatures. Because stable-isotope differences between food sources were not reflected in differences between life-history stages, the authors conservatively concluded that there was insufficient evidence for a diet shift with age. The ontogenetic shifts in fatty-acid profiles were intriguing, but the authors suggested that these reflected age-related physiological changes rather than changes in diet.
The authors’ work highlights the need to consider potential changes in the roles of invasive species with age, especially when evaluating interactions with native species. In this case, C. fornicata consumed a variety of food sources, including both benthic and particulate organic matter, regardless of age. The carbon stable-isotope signature of C. fornicata overlaps with those of several native suspension- and deposit-feeding species in the region [7], suggesting the possibility of resource competition, especially given the high abundances of this invader. This contribution demonstrates the potential difficulty of characterizing the impacts of an abundant invasive species with a complex life-history strategy. Like many invasive species, C. fornicata appears to be a dietary generalist, which likely contributes to its success in establishing and thriving in a variety of locations [8].
References
[1] Blanchard M (1997) Spread of the slipper limpet Crepidula fornicata (L. 1758) in Europe. Current state dans consequences. Scientia Marina, 61, 109–118. Open Access version : https://archimer.ifremer.fr/doc/00423/53398/54271.pdf
[2] Martin S, Thouzeau G, Chauvaud L, Jean F, Guérin L, Clavier J (2006) Respiration, calcification, and excretion of the invasive slipper limpet, Crepidula fornicata L.: Implications for carbon, carbonate, and nitrogen fluxes in affected areas. Limnology and Oceanography, 51, 1996–2007. https://doi.org/10.4319/lo.2006.51.5.1996
[3] Navarro JM, Chaparro OR (2002) Grazing–filtration as feeding mechanisms in motile specimens of Crepidula fecunda (Gastropoda: Calyptraeidae). Journal of Experimental Marine Biology and Ecology, 270, 111–122. https://doi.org/10.1016/S0022-0981(02)00013-8
[4] Yee AK, Padilla DK (2015) Allometric Scaling of the Radula in the Atlantic Slippersnail Crepidula fornicata. Journal of Shellfish Research, 34, 903–907. https://doi.org/10.2983/035.034.0320
[5] Collin R (1995) Sex, Size, and Position: A Test of Models Predicting Size at Sex Change in the Protandrous Gastropod Crepidula fornicata. The American Naturalist, 146, 815–831. https://doi.org/10.1086/285826
[6] Androuin T, Dubois SF, Hubas C, Lefebvre G, Grand FL, Schaal G, Carlier A (2021) Trophic niche of the invasive gregarious species Crepidula fornicata, in relation to ontogenic changes. bioRxiv, 2020.07.30.229021, ver. 4 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2020.07.30.229021
[7] Dauby P, Khomsi A, Bouquegneau J-M (1998) Trophic Relationships within Intertidal Communities of the Brittany Coasts: A Stable Carbon Isotope Analysis. Journal of Coastal Research, 14, 1202–1212. Retrieved May 4, 2021, from http://www.jstor.org/stable/4298880
[8] Machovsky-Capuska GE, Senior AM, Simpson SJ, Raubenheimer D (2016) The Multidimensional Nutritional Niche. Trends in Ecology & Evolution, 31, 355–365. https://doi.org/10.1016/j.tree.2016.02.009
| Trophic niche of the invasive gregarious species Crepidula fornicata, in relation to ontogenic changes | Thibault Androuin, Stanislas F. Dubois, Cédric Hubas, Gwendoline Lefebvre, Fabienne Le Grand, Gauthier Schaal, Antoine Carlier | <p style="text-align: justify;">The slipper limpet Crepidula fornicata is a common and widespread invasive gregarious species along the European coast. Among its life-history traits, well-documented ontogenic changes in behavior (i.e., motile male... |  | Food webs, Life history, Marine ecology | Matthew Bracken | | 2020-08-01 23:55:57 | View |
Consistent variations in personality traits and their potential for genetic improvement of biocontrol agents: Trichogramma evanescens as a case study
Silène Lartigue, Myriam Yalaoui, Jean Belliard, Claire Caravel, Louise Jeandroz, Géraldine Groussier, Vincent Calcagno, Philippe Louâpre, François-Xavier Dechaume-Moncharmont, Thibaut Malausa and Jérôme Moreau
https://doi.org/10.1101/2020.08.21.257881
Tell us how you can be, and we’ll make you better: exploiting genetic variability in personality traits to improve top-down control of agricultural pests
Recommended by Marta Montserrat based on reviews by Bart A Pannebakker, François Dumont, Joshua Patrick Byrne and Ana Pimenta Goncalves Pereira
Agriculture in the XXI century faces the huge challenge of having to provide food to a rapidly growing human population, which is expected to reach 10.9 billion in 2100 (UUNN 2019), by means of practices and methods that guarantee crop sustainability, human health safety, and respect to the environment (UUNN 2015). Such regulation by the United Nations ultimately entails that agricultural scientists are urged to design strategies and methods that effectively minimize the use of harmful chemical products to control pest populations and to improve soil quality.
One of the most, if not the most, sustainable, safe, and environmentally friendly approach to apply against pests is Biological Pest Control (BPC, hereafter), that is, the use of natural enemies to control the populations of pest organisms. The concept of BPC is by no means new: long back to the 300 AC, Chinese farmers built bamboo bridges between citrus trees to facilitate the foraging of the ant species Oecophylla smaragdina to control lepidopteran citrus pests (Konishi and Ito, 1973); It is also nice to use this recommendation letter to recall and quote the words written in 1752 by the famous Swedish taxonomist, botanist and zoologist, Carl Linnaeus: "Every insect has its predator which follows and destroys it. Such predatory insects should be caught and used for disinfecting crop-plants" (Hörstadius (1974) apud Linnaeus 1752).
Acknowledging the many cases of successes from BPC along our recent history, it is also true that application of BPC strategies during the XX century suffered from wrong-doings, mainly when the introduced biological control agent (BCA, hereafter) was of exotic origin and with a generalist diet-breath; in some cases the release of exotic species resulted on global extinction, reduction in the range of distribution, reduction in the population abundance, and partial displacement, of native and functionally similar species, and interbreeding with them (reviewed in van Lenteren et al. 2006). One of the most famous cases is that of Harmonia axyridis, a coccinellid predator of Asian origin that caused important environmental damage in North America (reviewed in Koch & Galvan, 2008).
Fortunately, after the implementation of the Nagoya protocol (CBD, 2011) importation of exotic species for BPC use was severely restricted and controlled, worldwide. Consequently, companies and agricultural scientist were driven to reinforce their focus and interest on the exploitation of native natural enemies, via the mass-rearing and release of native candidates (augmentative BPC), the conservation of landscapes near the crops to provide resources for natural enemies (i.e. conservation biological pest control), or via the exploitation of the genetic variability of BCAs, to create strains performing better at regulating pest populations under specific biotic or abiotic negative circumstances. Some of these cases are cited in Lartigue et al. (2020). The genetic improvement of BCAs is a strategy still in its infancy, but there is no doubt that the interest for it has significantly increased over the last 5 years (Lommen et al 2017, Bielza 2020, Leung et al 2020).
In my humble opinion, what makes the paper of Lartigue et al. (2020) a remarkable contribution to the field of genetic breeding of BCAs is that it opens a new window of opportunities to the field, by exploring the possibilities for artificial selection of behavioral traits (Réale et al. 2007) to "create" strains of natural enemies displaying behavioral syndromes (Sih et al. 2004) that makes them better at regulating pest populations. The behavioral approach for breeding BCAs can then be extended by crossing it with known abiotic and/or biotic hostile environments (e.g. warm and drought environments, presence of predators/competitors to the BCA, respectively) and engineer strains more prompt to display particular behavioral syndromes to help them to overcome the overall hostility of specific environments. I strongly believe that the approach proposed in Lartigue et al. (2020) will influence the future management of agricultural systems, where strategies including the genetic breeding of BCAs’ behavior will contribute to create better guards and protectors of our crops.
References
Bielza, P., Balanza, V., Cifuentes, D. and Mendoza, J. E. (2020). Challenges facing arthropod biological control: Identifying traits for genetic improvement of predators in protected crops. Pest Manag Sci. doi: https://doi.org/10.1002/ps.5857
CBD - Convention on Biological Diversity, 2011. The Nagoya Protocol on Access and Benefit-sharing, https://www.cbd.int/abs/doc/protocol/nagoya-protocol-en.pdf
Hörstadius, S. (1974). Linnaeus, animals and man. Biological Journal of the Linnaean Society, 6, 269-275. doi: https://doi.org/10.1111/j.1095-8312.1974.tb00725.x
Koch, R.L. and Galvan, T.L. (2008). Bad side of a good beetle: the North American experience with Harmonia axyridis. BioControl 53, 23–35. doi: https://doi.org/10.1007/978-1-4020-6939-0_3
Konishi, M. and Ito, Y. (1973). Early entomology in East Asia. In: Smith, R.F., Mittler, T.E., Smith, C.N. (Eds.), History of Entomology, Annual Reviews Inc., Palo Alto, California, pp. 1-20.
Lartigue, S., Yalaoui, M., Belliard, J., Caravel, C., Jeandroz, L., Groussier, G., Calcagno, V., Louâpre, P., Dechaume-Moncharmont, F.-X., Malausa, T. and Moreau, J. (2020). Consistent variations in personality traits and their potential for genetic improvement of biocontrol agents: Trichogramma evanescens as a case study. bioRxiv, 2020.08.21.257881, ver. 4 peer-reviewed and recommended by PCI Ecology. doi: https://doi.org/10.1101/2020.08.21.257881
Leung et al. (2020). Next-generation biological control: the need for integrating genetics and genomics. Biological Reviews, 95(6), 1838–1854. doi: https://doi.org/10.1111/brv.12641
Lommen, S. T. E., de Jong, P. W. and Pannebakker, B. A. (2017). It is time to bridge the gap between exploring and exploiting: prospects for utilizing intraspecific genetic variation to optimize arthropods for augmentative pest control – a review. Entomologia Experimentalis et Applicata, 162: 108-123. doi: https://doi.org/10.1111/eea.12510
Réale, D., Reader, S. M., Sol, D., McDougall, P. T. and Dingemanse, N. J. (2007). Integrating animal temperament within ecology and evolution. Biological Reviews, 82: 291-318. doi: https://doi.org/10.1111/j.1469-185X.2007.00010.x
Sih, A., Bell, A. and Johnson, J. C. (2004). Behavioral syndromes: an ecological and evolutionary overview. Trends in Ecology and Evolution, 19(7), 372–378. doi: https://doi.org/10.1016/j.tree.2004.04.009
UUNN. 2015. Transforming our world: the 2030 Agenda for Sustainable Development. report of the Open Working Group of the General Assembly on Sustainable Development Goals (A/68/970 and Corr.1; see also A/68/970/Add.1–3).
UUNN. 2019. World population prospects 2019. United Nations, Department of Economic and Social Affairs, Population Division: Highlights. ST/ESA/SER.A/423.
van Lenteren, J. C., Bale, J., Bigler, F., Hokkanen, H. M. T. and Loomans A. J. M. (2006). Assessing risks of releasing exotic biological control agents of arthropod pests. Annual Review of Entomology, 51: 609-634. doi: https://doi.org/10.1146/annurev.ento.51.110104.151129
| Consistent variations in personality traits and their potential for genetic improvement of biocontrol agents: Trichogramma evanescens as a case study | Silène Lartigue, Myriam Yalaoui, Jean Belliard, Claire Caravel, Louise Jeandroz, Géraldine Groussier, Vincent Calcagno, Philippe Louâpre, François-Xavier Dechaume-Moncharmont, Thibaut Malausa and Jérôme Moreau | <p>Improvements in the biological control of agricultural pests require improvements in the phenotyping methods used by practitioners to select efficient biological control agent (BCA) populations in industrial rearing or field conditions. Consist... |  | Agroecology, Behaviour & Ethology, Biological control, Evolutionary ecology, Life history | Marta Montserrat | | 2020-08-24 10:40:03 | View |
Body temperatures, life history, and skeletal morphology in the nine-banded armadillo (Dasypus novemcinctus)
Frank Knight, Cristin Connor, Ramji Venkataramanan, Robert J. Asher
https://doi.org/10.17863/CAM.50971
Is vertebral count in mammals influenced by developmental temperature? A study with Dasypus novemcinctus
Recommended by Mar Sobral based on reviews by Darin Croft and ?
Mammals show a very low level of variation in vertebral count, both among and within species, in comparison to other vertebrates [1]. Jordan’s rule for fishes states that the vertebral number among species increases with latitude, due to ambient temperatures during development [2]. Temperature has also been shown to influence vertebral count within species in fish [3], amphibians [4], and birds [5]. However, in mammals the count appears to be constrained, on the one hand, by a possible relationship between the development of the skeleton and the proliferations of cell lines with associated costs (neural malformations, cancer etc., [6]), and on the other by the cervical origin of the diaphragm [7].
Knight et al. [8] investigate the effect of intrauterine temperature variation on skeletal morphology during development, and focus on a particular mammal, Dasypus novemcinctus, or nine-banded armadillo. Armadillos (Xenarthra) and are characterized by relatively low body temperatures and low basal rates of metabolism. Dasypus novemcinctus is the only xenarthran mammal to have naturally expanded its range into the middle latitudes of the U.S., and one of the few mammals that invaded North America from South America. It is one of few placentals that withstand considerable decrease of body temperature without torpor. It presents a resting body temperature that is low and variable for a placental mammal of its size [9] and is the only vertebrate that gives birth to monozygotic quadruplets. Among 42 monotreme, marsupial and placental genera, Dasypus novemcinctus shows the highest variation of thoracolumbar vertebral count [10].
The particularities of Dasypus novemcinctus regarding vertebral count variation and ability to withstand variable temperature qualify it as a target organism for study of the relationship between skeleton morphology and temperature in mammals.
Knight et al. [8] explored variability in vertebral count within Dasypus novemcinctus to understand whether temperature during development determines skeleton morphology. To this end they experimented with 22 armadillos (19 with data) and litters from 12 pregnant females, in two environments, for three years — an impressive effort and experimental setup. Moreover, they used a wide variety of advanced experimental and analytical techniques. For example, they implanted intra-abdominal, long-term temperature recorders, which recorded data every 6 to 120 minutes for up to several months. They analysed body temperature periodicity by approximation of the recordings with Fourier series, and they CT-scanned fetuses.
All 19 individuals (from which data could be gathered) exhibited substantial daily variation in body temperature. Several intriguing results emerged such as the counter-intuitive finding that the mammals’ body temperature fluctuates more indoors than outdoors. Furthermore, three females (out of 12) were found to have offspring with atypical skeletons, and two of these mothers presented an extremely low internal temperature early in pregnancy. Additionally, genetically identical quadruplets differed skeletally among themselves within two litters.
Results are not yet definitive about the relationship of temperature during development and vertebral count in Dasypus novemcinctus. However, Knight et al. [8] demonstrated that nine-banded armadillos survive with high daily internal temperature fluctuations and successfully bring to term offspring which vary in skeletal morphology among and within genetically identical litters despite major temperature extremes.
References
[1] Hautier L, Weisbecker V, Sánchez-Villagra MR, Goswami A, Asher RJ (2010) Skeletal development in sloths and the evolution of mammalian vertebral patterning. Proceedings of the National Academy of Sciences, 107, 18903–18908. doi: 10.1073/pnas.1010335107
[2] Jordan, D.S. (1892) Relations of temperature to vertebrae among fishes. Proceedings of the United States National Museum, 1891, 107-120. doi: 10.5479/si.00963801.14-845.107
[3] Tibblin P, Berggren H, Nordahl O, Larsson P, Forsman A (2016) Causes and consequences of intra-specific variation in vertebral number. Scientific Reports, 6, 1–12. doi: 10.1038/srep26372
[4] Peabody RB, Brodie ED (1975) Effect of temperature, salinity and photoperiod on the number of trunk vertebrae in Ambystoma maculatum. Copeia, 1975, 741–746. doi: 10.2307/1443326
[5] Lindsey CC, Moodie GEE (1967) The effect of incubation temperature on vertebral count in the chicken. Canadian Journal of Zoology, 45, 891–892. doi: 10.1139/z67-099
[6] Galis F, Dooren TJMV, Feuth JD, Metz JAJ, Witkam A, Ruinard S, Steigenga MJ, Wunaendts LCD (2006) Extreme selection in humans against homeotic transformations of cervical vertebrae. Evolution, 60, 2643–2654. doi: 10.1111/j.0014-3820.2006.tb01896.x
[7] Buchholtz EA, Stepien CC (2009) Anatomical transformation in mammals: developmental origin of aberrant cervical anatomy in tree sloths. Evolution and Development, 11, 69–79. doi: 10.1111/j.1525-142X.2008.00303.x
[8] Knight F, Connor C, Venkataramanan R, Asher RJ. (2020). Body temperatures, life history, and skeletal morphology in the nine-banded armadillo (Dasypus novemcinctus). PCI-Ecology. doi: 10.17863/CAM.50971
[9] McNab BK (1980) Energetics and the limits to a temperate distribution in armadillos. Journal of Mammalogy, 61, 606–627. doi: 10.2307/1380307
[10] Asher RJ, Lin KH, Kardjilov N, Hautier L (2011) Variability and constraint in the mammalian vertebral column. Journal of Evolutionary Biology, 24, 1080–1090. doi: 10.1111/j.1420-9101.2011.02240.x
| Body temperatures, life history, and skeletal morphology in the nine-banded armadillo (Dasypus novemcinctus) | Frank Knight, Cristin Connor, Ramji Venkataramanan, Robert J. Asher | <p>The nine banded armadillo (*Dasypus novemcinctus*) is the only xenarthran mammal to have naturally expanded its range into the middle latitudes of the USA. It is not known to hibernate, but has been associated with unusually labile core body te... |  | Behaviour & Ethology, Evolutionary ecology, Life history, Physiology, Zoology | Mar Sobral | | 2019-11-22 22:57:31 | View |
Studying a rare behavior in a polygamous bird: male parental care in great-tailed grackles
Recommended by Marie-Jeanne Holveck based on reviews by Matthieu Paquet and André C Ferreira
The Great-tailed grackle (Quiscalus mexicanus) is a polygamous bird species that is originating from Central America and rapidly expanding its geographic range toward the North, and in which females were long thought to be the sole nest builders and caretakers of the young. In their pre-registration [1], Folsom and collaborators report repeated occurrences of male parental care and develop hypotheses aiming at better understanding the occurrence and the fitness consequences of this very rarely observed male behavior. They propose to assess if male parental care correlates with the circulating levels of several relevant hormones, increases offspring survival, is a local adaptation, and is a mating strategy, in surveying three populations located in Arizona (middle of the geographic range expansion), California (northern edge of the geographic range), and in Central America (core of the range). This study is part of a 5-year bigger project.
Both reviewers and I strongly value Folsom and collaborators’ commitment to program a study, in natural field conditions, of a rare, yet likely evolutionary-important behavior, namely parental care by males of the great-tailed grackle. Yet, we all also recognized that it is a risky endeavor, and as a consequence, we wondered about the authors’ ability to reach a sufficient sample size to statistically test (all) hypotheses and predictions with enough confidence (e.g. risk of type I errors, also known as false positives).
Folsom and collaborators acknowledged these limitations in their pre-registration. (i) They made the exploratory nature of their research work clear to readers. (ii) They adapted their analysis plan in running prior power analyses and in focusing on effect sizes (estimates and confidence intervals). (iii) Last and not least, Folsom and collaborators clearly exposed a priori hypotheses, their associated predictions and alternatives, and ranked the latter based on their plausibility according to knowledge in the current and other study systems. Developing theory about male parental care behavior more generally with regard to a polygamous species that is rapidly expanding its geographic range and that is considered not to provide male parental care is without any doubt an added value to this study.
In summary, while this study will likely be insufficient to fully understand male parental care behavior of great-tailed grackles, it is much needed because it will definitely allow rejecting some predictions (e.g., if this behavior is present in all the studied populations, it would be common across range against expectation; finding only one male providing care to an unrelated offspring would lead to reject the prediction that males only care for their own offspring) and thus it will help laying the foundation of future research directions.
I strongly support the pre-registration system and thank all the contributors for producing a fruitful discussion throughout the review process, which in fine improved the clarity and logic of this pre-registration. Given the positive and encouraging reviews, the detailed and thorough answers to all comments by Folsom and collaborators, and their satisfactory and interesting revisions, I am happy to recommend this pre-registration and I look forward to seeing its outcomes.
References
[1] Folsom MA, MacPherson M, Lukas D, McCune KB, Bergeron L, Bond A, Blackwell A, Rowney C, Logan CJ. 2020. Investigating the rare behavior of male parental care in great-tailed grackles. corinalogan.com/Preregistrations/gmalecare.html In principle acceptance by PCI Ecology of the version on 15 June 2020 corinalogan/grackles/blob/master/Files/Preregistrations/gmalecare.Rmd.
| Investigating the rare behavior of male parental care in great-tailed grackles | Folsom MA, MacPherson M, Lukas D, McCune KB, Bergeron L, Bond A, Blackwell A, Rowney C, Logan CJ | This is a PREREGISTRATION submitted for pre-study peer review. Our planned data collection START DATE is May 2020, therefore it would be ideal if the peer review process could be completed before then.
Abstract: Great-tailed grackles (Quiscalus... |  | Behaviour & Ethology, Biological invasions, Preregistrations, Zoology | Marie-Jeanne Holveck | | 2019-12-05 17:38:47 | View |
Impact of group management and transfer on individual sociality in Highland cattle (Bos Taurus)
Sebastian Sosa, Marie Pelé, Elise Debergue, Cedric Kuntz, Blandine Keller, Florian Robic, Flora Siegwalt-Baudin, Camille Richer, Amandine Ramos, Cédric Sueur
https://arxiv.org/abs/1805.11553v4
How empirical sciences may improve livestock welfare and help their management
Recommended by Marie Charpentier based on reviews by Alecia CARTER and 1 anonymous reviewer
Understanding how livestock management is a source of social stress and disturbances for cattle is an important question with potential applications for animal welfare programs and sustainable development. In their article, Sosa and colleagues [1] first propose to evaluate the effects of individual characteristics on dyadic social relationships and on the social dynamics of four groups of cattle. Using network analyses, the authors provide an interesting and complete picture of dyadic interactions among groupmates. Although shown elsewhere, the authors demonstrate that individuals that are close in age and close in rank form stronger dyadic associations than other pairs. Second, the authors take advantage of some transfers of animals between groups -for management purposes- to assess how these transfers affect the social dynamics of groupmates. Their central finding is that the identity of transferred animals is a key-point. In particular, removing offspring strongly destabilizes the social relationships of mothers while adding a bull into a group also profoundly impacts female-female social relationships, as social networks before and after transfer of these key-animals are completely different. In addition, individuals, especially the young ones, that are transferred without familiar conspecifics take more time to socialize with their new group members than individuals transferred with familiar groupmates, generating a potential source of stress. Interestingly, the authors end up their article with some thoughts on the implications of their findings for animal welfare and ethics. This study provides additional evidence that empirical science has a major role to play in providing recommendations regarding societal questions such as livestock management and animal wellbeing.
References
[1] Sosa, S., Pelé, M., Debergue, E., Kuntz, C., Keller, B., Robic, F., Siegwalt-Baudin, F., Richer, C., Ramos, A., & Sueur C. (2018). Impact of group management and transfer on individual sociality in Highland cattle (Bos Taurus). arXiv:1805.11553v4 [q-bio.PE] peer-reviewed and recommended by PCI Ecol. https://arxiv.org/abs/1805.11553v4
| Impact of group management and transfer on individual sociality in Highland cattle (Bos Taurus) | Sebastian Sosa, Marie Pelé, Elise Debergue, Cedric Kuntz, Blandine Keller, Florian Robic, Flora Siegwalt-Baudin, Camille Richer, Amandine Ramos, Cédric Sueur | The sociality of cattle facilitates the maintenance of herd cohesion and synchronisation, making these species the ideal choice for domestication as livestock for humans. However, livestock populations are not self-regulated, and farmers transfer ... |  | Behaviour & Ethology, Social structure | Marie Charpentier | | 2018-05-30 14:05:39 | View |
Are condition indices positively related to each other and to fitness?: a test with grackles
Recommended by Marcos Mendez based on reviews by Javier Seoane and Isabel López-Rull
Reproductive succes, as a surrogate of individual fitness, depends both on extrinsic and intrinsic factors [1]. Among the intrinsic factors, resource level or health are considered important potential drivers of fitness but exceedingly difficult to measure directly. Thus, a host of proxies have been suggested, known as condition indices [2]. The question arises whether all condition indices consistently measure the same "inner state" of individuals and whether all of them similarly correlate to individual fitness. In this preregistration, Berens and colleagues aim to answer this question for two common condition indices, fat score and scaled mass index (Fig. 1), using great-tailed grackles as a model system. Although this question is not new, it has not been satisfactorily solved and both reviewers found merit in the attempt to clarify this matter.

Figure 1. Hypothesized relationships between two condition indices and reproductive success. Single arrow heads indicate causal relationships; double arrow heads indicate only correlation. In a best case scenario, all relationships should be positive and linear.
A problem in adressing this question with grackles is limited population, ergo sample, size and limited possibilites of recapture individuals. Some relationships can be missed due to low statistical power. Unfortunately, existing tools for power analysis fall behind complex designs and the one planned for this study. Thus, any potentially non significant relationship has to be taken cautiously. Nevertheless, even if grackles will not provide a definitive answer (they never meant to do it), this preregistration can inspire broader explorations of matches and mismatches across condition indices and species, as well as uncover non-linear relationships with reproductive success.
References
[1] Roff, D. A. (2001). Life history evolution. Oxford University Press, Oxford.
[2] Labocha, M. K.; Hayes, J. P. (2012). Morphometric indices of body condition in birds: a review. Journal of Ornithology 153: 1–22. doi: 10.1007/s10336-011-0706-1
| Validating morphological condition indices and their relationship with reproductive success in great-tailed grackles | Jennifer M. Berens, Corina J. Logan, Melissa Folsom, Luisa Bergeron, Kelsey B. McCune | Morphological variation among individuals has the potential to influence multiple life history characteristics such as dispersal, migration, reproductive fitness, and survival (Wilder, Raubenheimer, and Simpson (2016)). Theoretically, individuals ... |  | Behaviour & Ethology, Conservation biology, Demography, Morphometrics, Preregistrations, Zoology | Marcos Mendez | | 2019-08-05 20:05:56 | View |
Recommendations to address uncertainties in environmental risk assessment using toxicokinetics-toxicodynamics models
Virgile Baudrot and Sandrine Charles
https://doi.org/10.1101/356469
Addressing uncertainty in Environmental Risk Assessment using mechanistic toxicological models coupled with Bayesian inference
Recommended by Luis Schiesari based on reviews by Andreas Focks and 2 anonymous reviewers
Environmental Risk Assessment (ERA) is a strategic conceptual framework to characterize the nature and magnitude of risks, to humans and biodiversity, of the release of chemical contaminants in the environment. Several measures have been suggested to enhance the science and application of ERA, including the identification and acknowledgment of uncertainties that potentially influence the outcome of risk assessments, and the appropriate consideration of temporal scale and its linkage to assessment endpoints [1].
Baudrot & Charles [2] proposed to approach these questions by coupling toxicokinetics-toxicodynamics models, which describe the time-course of processes leading to the adverse effects of a toxicant, with Bayesian inference. TKTD models separate processes influencing an organismal internal exposure (´toxicokinetics´, i.e., the uptake, bioaccumulation, distribution, biotransformation and elimination of a toxicant) from processes leading to adverse effects and ultimately its death (´toxicodynamics´) [3]. Although species and substance specific, the mechanistic nature of TKTD models facilitates the comparison of different toxicants, species, life stages, environmental conditions and endpoints [4].
Baudrot & Charles [2] investigated the use of a Bayesian framework to assess the uncertainties surrounding the calibration of General Unified Threshold Models of Survival (a category of TKTD) with data from standard toxicity tests, and their propagation to predictions of regulatory toxicity endpoints such as LC(x,t) [the lethal concentration affecting any x% of the population at any given exposure duration of time t] and MF(x,t) [an exposure multiplication factor leading to any x% effect reduction due to the contaminant at any time t].
Once calibrated with empirical data, GUTS models were used to explore individual survival over time, and under untested exposure conditions. Lethal concentrations displayed a strong curvilinear decline with time of exposure. For a given total amount of contaminant, pulses separated by short time intervals yielded higher mortality than pulses separated by long time intervals, as did few pulses of high amplitude when compared to multiple pulses of low amplitude. The response to a pulsed contaminant exposure was strongly influenced by contaminant depuration times. These findings highlight one important contribution of TKTD modelling in ecotoxicology: they represent just a few of the hundreds of exposure scenarios that could be mathematically explored, and that would be unfeasible or even unethical to conduct experimentally.
GUTS models were also used for interpolations or extrapolations of assessment endpoints, and their marginal distributions. A case in point is the incipient lethal concentration. The responses of model organisms to contaminants in standard toxicity tests are typically assessed at fixed times of exposure (e.g. 24h or 48h in the Daphnia magna acute toxicity test). However, because lethal concentrations are strongly time-dependent, it has been suggested that a more meaningful endpoint would be the incipient (i.e. asymptotic) lethal concentration when time of exposure increases to infinity. The authors present a mathematical solution for calculating the marginal distribution of such incipient lethal concentration, thereby providing both more relevant information and a way of comparing experiments, compounds or species tested for different periods of time.
Uncertainties were found to change drastically with time of exposure, being maximal at extreme values of x for both LC(x,t) and MF(x,t). In practice this means that assessment endpoints estimated when the effects of the contaminant are weak (such as LC10, the contaminant concentration resulting in the mortality of 10% of the experimental population), a commonly used assessment value in ERA, are prone to be highly variable.
The authors end with recommendations for improved experimental design, including (i) using assessment endpoints at intermediate values of x (e.g., LC50 instead of LC10) (ii) prolonging exposure and recording mortality over the course of the experiment (iii) experimenting one or few peaks of high amplitude close to each other when assessing pulsed exposure. Whereas these recommendations are not that different from current practices, they are based on a more coherent mechanistic grounding.
Overall, this and other contributions from Charles, Baudrot and their research group contribute to turn TKTD models into a real tool for Environmental Risk Assessment. Further enhancement of ERA´s science and application could be achieved by extending the use of TKTD models to sublethal rather than lethal effects, and to chronic rather than acute exposure, as these are more controversial issues in decision-making regarding contaminated sites.
References
[1] Dale, V. H., Biddinger, G. R., Newman, M. C., Oris, J. T., Suter, G. W., Thompson, T., ... & Chapman, P. M. (2008). Enhancing the ecological risk assessment process. Integrated environmental assessment and management, 4(3), 306-313. doi: 10.1897/IEAM_2007-066.1
[2] Baudrot, V., & Charles, S. (2018). Recommendations to address uncertainties in environmental risk assessment using toxicokinetics-toxicodynamics models. bioRxiv, 356469, ver. 3 peer-reviewed and recommended by PCI Ecol. doi: 10.1101/356469
[3] EFSA Panel on Plant Protection Products and their Residues (PPR), Ockleford, C., Adriaanse, P., Berny, P., Brock, T., Duquesne, S., ... & Kuhl, T. (2018). Scientific Opinion on the state of the art of Toxicokinetic/Toxicodynamic (TKTD) effect models for regulatory risk assessment of pesticides for aquatic organisms. EFSA Journal, 16(8), e05377. doi: 10.2903/j.efsa.2018.5377
[4] Jager, T., Albert, C., Preuss, T. G., & Ashauer, R. (2011). General unified threshold model of survival-a toxicokinetic-toxicodynamic framework for ecotoxicology. Environmental science & technology, 45(7), 2529-2540. doi: 10.1021/es103092a
| Recommendations to address uncertainties in environmental risk assessment using toxicokinetics-toxicodynamics models | Virgile Baudrot and Sandrine Charles | <p>Providing reliable environmental quality standards (EQS) is a challenging issue for environmental risk assessment (ERA). These EQS are derived from toxicity endpoints estimated from dose-response models to identify and characterize the environm... | 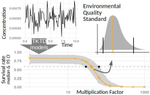 | Chemical ecology, Ecotoxicology, Experimental ecology, Statistical ecology | Luis Schiesari | | 2018-06-27 21:33:30 | View |
Parasites make hosts more profitable but less available to predators
Loïc Prosnier, Nicolas Loeuille, Florence D. Hulot, David Renault, Christophe Piscart, Baptiste Bicocchi, Muriel Deparis, Matthieu Lam, Vincent Médoc
https://doi.org/10.1101/2022.02.08.479552
Indirect effects of parasitism include increased profitability of prey to optimal foragers
Recommended by Luis Schiesari based on reviews by Thierry DE MEEUS and Eglantine Mathieu-Bégné
Even though all living organisms are, at the same time, involved in host-parasite interactions and embedded in complex food webs, the indirect effects of parasitism are only beginning to be unveiled.
Prosnier et al. investigated the direct and indirect effects of parasitism making use of a very interesting biological system comprising the freshwater zooplankton Daphnia magna and its highly specific parasite, the iridovirus DIV-1 (Daphnia-iridescent virus 1). Daphnia are typically semitransparent, but once infected develop a white phenotype with a characteristic iridescent shine due to the enlargement of white fat cells.
In a combination of infection trials and comparison of white and non-white phenotypes collected in natural ponds, the authors demonstrated increased mortality and reduced lifetime fitness in infected Daphnia. Furthermore, white phenotypes had lower mobility, increased reflectance, larger body sizes and higher protein content than non-white phenotypes. As a consequence, total energy content was effectively doubled in white Daphnia when compared to non-white broodless Daphnia.
Next the authors conducted foraging trials with Daphnia predators Notonecta (the backswimmer) and Phoxinus (the European minnow). Focusing on Notonecta, unchanged search time and increased handling time were more than compensated by the increased energy content of white Daphnia. White Daphnia were 24% more profitable and consistently preferred by Notonecta, as the optimal foraging theory would predict. The authors argue that menu decisions of optimal foragers in the field might be different, however, as the prevalence – and therefore availability - of white phenotypes in natural populations is very low.
The study therefore contributes to our understanding of the trophic context of parasitism. One shortcoming of the study is that the authors rely exclusively on phenotypic signs for determining infection. On their side, DIV-1 is currently known to be highly specific to Daphnia, their study site is well within DIV-1 distributional range, and the symptoms of infection are very conspicuous. Furthermore, the infection trial – in which non-white Daphnia were exposed to white Daphnia homogenates - effectively caused several lethal and sublethal effects associated with DIV-1 infection, including iridescence. However, the infection trial also demonstrated that part of the exposed individuals developed intermediate traits while still keeping the non-white, non-iridescent phenotype. Thus, there may be more subtleties to the association of DIV-1 infection of Daphnia with ecological and evolutionary consequences, such as costs to resistance or covert infection, that the authors acknowledge, and that would be benefitted by coupling experimental and observational studies with the determination of actual infection and viral loads.
References
Prosnier L., N. Loeuille, F.D. Hulot, D. Renault, C. Piscart, B. Bicocchi, M, Deparis, M. Lam, & V. Médoc. (2023). Parasites make hosts more profitable but less available to predators. BioRxiv, ver. 4 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2022.02.08.479552
| Parasites make hosts more profitable but less available to predators | Loïc Prosnier, Nicolas Loeuille, Florence D. Hulot, David Renault, Christophe Piscart, Baptiste Bicocchi, Muriel Deparis, Matthieu Lam, Vincent Médoc | <p>Parasites are omnipresent, and their eco-evolutionary significance has aroused much interest from scientists. Parasites may affect their hosts in many ways by altering host density, vulnerability to predation, and energy content, thus modifying... |  | Community ecology, Eco-evolutionary dynamics, Epidemiology, Experimental ecology, Food webs, Foraging, Freshwater ecology, Host-parasite interactions, Life history, Parasitology, Statistical ecology | Luis Schiesari | | 2022-05-20 10:15:41 | View |
Comparing statistical and mechanistic models to identify the drivers of mortality within a rear-edge beech population
Cathleen Petit-Cailleux, Hendrik Davi, François Lefevre, Christophe Hurson, Joseph Garrigue, Jean-André Magdalou, Elodie Magnanou and Sylvie Oddou-Muratorio
https://doi.org/10.1101/645747
The complexity of predicting mortality in trees
Recommended by Lucía DeSoto based on reviews by Lisa Hülsmann and 2 anonymous reviewers
One of the main issues of forest ecosystems is rising tree mortality as a result of extreme weather events (Franklin et al., 1987). Eventually, tree mortality reduces forest biomass (Allen et al., 2010), although its effect on forest ecosystem fluxes seems not lasting too long (Anderegg et al., 2016). This controversy about the negative consequences of tree mortality is joined to the debate about the drivers triggering and the mechanisms accelerating tree decline. For instance, there is still room for discussion about carbon starvation or hydraulic failure determining the decay processes (Sevanto et al., 2014) or about the importance of mortality sources (Reichstein et al., 2013). Therefore, understanding and predicting tree mortality has become one of the challenges for forest ecologists in the last decade, doubling the rate of articles published on the topic (*). Although predicting the responses of ecosystems to environmental change based on the traits of species may seem a simplistic conception of ecosystem functioning (Sutherland et al., 2013), identifying those traits that are involved in the proneness of a tree to die would help to predict how forests will respond to climate threatens.
Modelling tree mortality is complex, involving multiple factors acting simultaneously at different scales, from tree genetics to ecosystem dynamics and from microsite conditions to global climatic events. Therefore, taking into account different approaches to reduce uncertainty of the predictions is needed (Bugmann et al., 2019). Petit-Cailleux et al. (2020) uses statistical and process-based models to detect the main mortality drivers of a drought- and frost-prone beech population. Particularly, they assessed the intra-individual characteristics of the population, that may play a decisive role explaining the differences in tree vulnerability to extreme weather events. Comparing the results of both analytical approaches, they find out several key factors, such as defoliation, leaf phenology and tree size, that were consistent between them. Even more, the process-based model showed the physiological mechanisms that may explain the individual vulnerability, for instance higher loss of hydraulic conductance may increase the mortality risk of trees with early budburst phenology and large stem diameter. The authors also successfully model annual mortality rate with a linear relationship including only three parameters: loss of conductance, biomass of reserves and late frost days.
This valuable study is a good example of the complexity in understanding and predicting tree mortality. The authors carried out the ambitious commitment of studying the inter-annual variation in mortality with 14-year dataset. However, it might be not enough time to control for the dependence of temporal data to soundly model mortality rate. The authors also acknowledge that the use of two approaches increases the knowledge from different perspectives, but at the same time comparing their results is difficult because the parameters used are not identical. Particularly, process-based models tend to consider the same microclimatic conditions for every tree in the population, and may produce inconsistences with statistical models. Alternatively, individual-based modelling might overcome some of the incompatibilities between the approaches (Zhu et al., 2019).
(*) Number (and percentage) of articles found in Web of Sciences after searching (December the 10th, 2020) “tree mortality”: from 163 (0.006%) in 2010 to 412 (0.013%) in 2020.
References
Allen et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest ecology and management, 259(4), 660-684. doi: https://doi.org/10.1016/j.foreco.2009.09.001
Anderegg et al. (2016). When a tree dies in the forest: scaling climate-driven tree mortality to ecosystem water and carbon fluxes. Ecosystems, 19(6), 1133-1147. doi: https://doi.org/10.1007/s10021-016-9982-1
Bugmann et al. (2019). Tree mortality submodels drive simulated long‐term forest dynamics: assessing 15 models from the stand to global scale. Ecosphere, 10(2), e02616. doi: https://doi.org/10.1002/ecs2.2616
Franklin, J. F., Shugart, H. H. and Harmon, M. E. (1987) Death as an ecological process: the causes, consequences, and variability of tree mortality. BioScience, 37, 550–556. doi: https://doi.org/10.2307/1310665
Petit-Cailleux, C., Davi, H., Lefèvre, F., Garrigue, J., Magdalou, J.-A., Hurson, C., Magnanou, E. and Oddou-Muratorio, S. (2020) Comparing statistical and mechanistic models to identify the drivers of mortality within a rear-edge beech population. bioRxiv, 645747, ver 7 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/645747
Reichstein et al. (2013). Climate extremes and the carbon cycle. Nature, 500(7462), 287-295. doi: https://doi.org/10.1038/nature12350
Sevanto, S., Mcdowell, N. G., Dickman, L. T., Pangle, R., and Pockman, W. T. (2014). How do trees die? A test of the hydraulic failure and carbon starvation hypotheses. Plant, cell & environment, 37(1), 153-161. doi: https://doi.org/10.1111/pce.12141
Sutherland et al. (2013). Identification of 100 fundamental ecological questions. Journal of ecology, 101(1), 58-67. doi: https://doi.org/10.1111/1365-2745.12025
Zhu, Y., Liu, Z., and Jin, G. (2019). Evaluating individual-based tree mortality modeling with temporal observation data collected from a large forest plot. Forest Ecology and Management, 450, 117496. doi: https://doi.org/10.1016/j.foreco.2019.117496
| Comparing statistical and mechanistic models to identify the drivers of mortality within a rear-edge beech population | Cathleen Petit-Cailleux, Hendrik Davi, François Lefevre, Christophe Hurson, Joseph Garrigue, Jean-André Magdalou, Elodie Magnanou and Sylvie Oddou-Muratorio | <p>Since several studies have been reporting an increase in the decline of forests, a major issue in ecology is to better understand and predict tree mortality. The interactions between the different factors and the physiological processes giving ... |  | Climate change, Physiology, Population ecology | Lucía DeSoto | | 2019-05-24 11:37:38 | View |
The inherent multidimensionality of temporal variability: How common and rare species shape stability patterns
Jean-François Arnoldi, Michel Loreau, Bart Haegeman
https://doi.org/10.1101/431296
Diversity-Stability and the Structure of Perturbations
Recommended by Kevin Cazelles and Kevin Shear McCann based on reviews by Frederic Barraquand and 1 anonymous reviewer and Kevin Shear McCann based on reviews by Frederic Barraquand and 1 anonymous reviewer
In his 1972 paper “Will a Large Complex System Be Stable?” [1], May challenges the idea that large communities are more stable than small ones. This was the beginning of a fundamental debate that still structures an entire research area in ecology: the diversity-stability debate [2]. The most salient strength of May’s work was to use a mathematical argument to refute an idea based on the observations that simple communities are less stable than large ones. Using the formalism of dynamical systems and a major results on the distribution of the eigen values for random matrices, May demonstrated that the addition of random interactions destabilizes ecological communities and thus, rich communities with a higher number of interactions should be less stable. But May also noted that his mathematical argument holds true only if ecological interactions are randomly distributed and thus concluded that this must not be true! This is how the contradiction between mathematics and empirical observations led to new developments in the study of ecological networks.
Since 1972, the theoretical corpus of ecology has advanced, building on the formalism of dynamical systems, ecologists have revealed that ecological interactions are indeed not randomly distributed [3,4], but general rules are still missing and we are far from understanding what determine the exact network topology of a given community. One promising avenue is to understand the relationship between different facets of the concept of stability [5,6]. Indeed, the classical approach to determine whether a system is stable is qualitative: if a system returns to its equilibrium when it is slightly moved away from it, then the system is considered stable. But there are several other aspects that are worth scrutinizing. For instance, when a system returns to its equilibrium, one can characterize the corresponding transient dynamics [7,8], that is asking fundamental questions such as: what is the trajectory of return? How long does it take to return to the equilibrium? Another fundamental question is whether the system remains qualitatively stable when the distributions of interactions strengths change? From a biological standpoint, all of these questions matter as all these aspects of stability may partially explain the actual structure of ecological networks, and hence, frameworks that integrate several facets of stability are much needed.
The study by Arnoldi et al. [9] is a significant step towards such a framework. The strength of their formalism is threefold. First, instead of considering separately the system and its perturbations, they considering the fluctuations of a perturbed ecological systems and thus, perturbations are parts of the ecological system. Second, they use of a broad definition of perturbation that encompasses the types of perturbations (whether the individual respond synchronously or not), their intensity and their direction (how the perturbations are correlated across species). Third, they quantify the instability of the system using variability which integrates the consequences of perturbations over the whole set of species of a community: such a measure is comparable across communities and accounts for the trivial effect of the perturbations on the system dynamics.
Using this framework, the authors show that interactions within a stable community leads to a general relationship between variability and the abundance of individually perturbed species: if individuals of species respond in synchrony to a perturbation, then the more abundant the species perturbed the higher the variability of the system, but the relationship is reverse when individual respond asynchronously. A direct implications of these results for the classical debate is that the diversity-stability relationship is negative for the former type of perturbations (as in May’s seminal paper) but positive for the latter type. Hence, the rigorous work of Arnoldi and colleagues sheds a new light upon the classical debate: the nature of the perturbation regime prevailing within a community affects the slope of the diversity-stability relationships and given the vast diversity of ecological communities, this may very well be one of the reasons why the debate still endures.
From a historical perspective, it is interesting that ecologists have gone from looking at random webs to structured webs and now, in a sense, Arnoldi et al. are unpacking the role of differentially structured perturbations. The work they achieved will doubtlessly be followed by further theoretical investigations. One natural research avenue is to revisit the role of the topology of ecological networks with this framework: how the distribution of interactions and their strength affect the general relationship they unravel? Finally, this study demonstrate that the impact of the abundance of a species on the variability of the system depends on the nature of the perturbation regime and so the distribution of species abundances within a community should be determined by the prevailing perturbation regime which is a prediction that remains to be tested.
References
[1] May, Robert M (1972). Will a Large Complex System Be Stable? Nature 238, 413–414. doi: 10.1038/238413a0
[2] McCann, Kevin Shear (2000). The Diversity–Stability Debate. Nature 405, 228–233. doi: 10.1038/35012234
[3] Rooney, Neil, Kevin McCann, Gabriel Gellner, and John C. Moore (2006). Structural Asymmetry and the Stability of Diverse Food Webs. Nature 442, 265–269. doi: 10.1038/nature04887
[4] Jacquet, Claire, Charlotte Moritz, Lyne Morissette, Pierre Legagneux, François Massol, Philippe Archambault, and Dominique Gravel (2016). No Complexity–Stability Relationship in Empirical Ecosystems. Nature Communications 7, 12573. doi: 10.1038/ncomms12573
[5] Donohue, Ian, Helmut Hillebrand, José M. Montoya, Owen L. Petchey, Stuart L. Pimm, Mike S. Fowler, Kevin Healy, et al. (2016). Navigating the Complexity of Ecological Stability. Ecology Letters 19, 1172–1185. doi: 10.1111/ele.12648
[6] Arnoldi, Jean-François, and Bart Haegeman (2016). Unifying Dynamical and Structural Stability of Equilibria. Proceedings of the Royal Society A: Mathematical, Physical and Engineering Science 472, 20150874. doi: 10.1098/rspa.2015.0874
[7] Caswell, Hal, and Michael G. Neubert (2005). Reactivity and Transient Dynamics of Discrete-Time Ecological Systems. Journal of Difference Equations and Applications 11, 295–310. doi: 10.1080/10236190412331335382
[8] Arnoldi, J-F., M. Loreau, and B. Haegeman (2016). Resilience, Reactivity and Variability: A Mathematical Comparison of Ecological Stability Measures. Journal of Theoretical Biology 389, 47–59. doi: 10.1016/j.jtbi.2015.10.012
[9] Arnoldi, Jean-Francois, Michel Loreau, and Bart Haegeman. (2019). The Inherent Multidimensionality of Temporal Variability: How Common and Rare Species Shape Stability Patterns.” BioRxiv, 431296, ver. 3 peer-reviewed and recommended by PCI Ecology. doi: 10.1101/431296
| The inherent multidimensionality of temporal variability: How common and rare species shape stability patterns | Jean-François Arnoldi, Michel Loreau, Bart Haegeman | <p>Empirical knowledge of ecosystem stability and diversity-stability relationships is mostly based on the analysis of temporal variability of population and ecosystem properties. Variability, however, often depends on external factors that act as... | 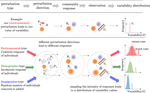 | Biodiversity, Coexistence, Community ecology, Competition, Interaction networks, Theoretical ecology | Kevin Cazelles | | 2018-10-02 14:01:03 | View |







