
SCHIESARI Luis
- Theoretical and Applied Ecology in Modified Environments, Universidade de São Paulo, São Paulo, Brazil
- Agroecology, Biodiversity, Community ecology, Conservation biology, Ecotoxicology, Experimental ecology, Freshwater ecology, Human impact, Spatial ecology, Metacommunities & Metapopulations, Tropical ecology
- recommender
Recommendations: 2
Reviews: 0
Recommendations: 2

Parasites make hosts more profitable but less available to predators
Indirect effects of parasitism include increased profitability of prey to optimal foragers
Recommended by Luis Schiesari based on reviews by Thierry DE MEEUS and Eglantine Mathieu-BégnéEven though all living organisms are, at the same time, involved in host-parasite interactions and embedded in complex food webs, the indirect effects of parasitism are only beginning to be unveiled.
Prosnier et al. investigated the direct and indirect effects of parasitism making use of a very interesting biological system comprising the freshwater zooplankton Daphnia magna and its highly specific parasite, the iridovirus DIV-1 (Daphnia-iridescent virus 1). Daphnia are typically semitransparent, but once infected develop a white phenotype with a characteristic iridescent shine due to the enlargement of white fat cells.
In a combination of infection trials and comparison of white and non-white phenotypes collected in natural ponds, the authors demonstrated increased mortality and reduced lifetime fitness in infected Daphnia. Furthermore, white phenotypes had lower mobility, increased reflectance, larger body sizes and higher protein content than non-white phenotypes. As a consequence, total energy content was effectively doubled in white Daphnia when compared to non-white broodless Daphnia.
Next the authors conducted foraging trials with Daphnia predators Notonecta (the backswimmer) and Phoxinus (the European minnow). Focusing on Notonecta, unchanged search time and increased handling time were more than compensated by the increased energy content of white Daphnia. White Daphnia were 24% more profitable and consistently preferred by Notonecta, as the optimal foraging theory would predict. The authors argue that menu decisions of optimal foragers in the field might be different, however, as the prevalence – and therefore availability - of white phenotypes in natural populations is very low.
The study therefore contributes to our understanding of the trophic context of parasitism. One shortcoming of the study is that the authors rely exclusively on phenotypic signs for determining infection. On their side, DIV-1 is currently known to be highly specific to Daphnia, their study site is well within DIV-1 distributional range, and the symptoms of infection are very conspicuous. Furthermore, the infection trial – in which non-white Daphnia were exposed to white Daphnia homogenates - effectively caused several lethal and sublethal effects associated with DIV-1 infection, including iridescence. However, the infection trial also demonstrated that part of the exposed individuals developed intermediate traits while still keeping the non-white, non-iridescent phenotype. Thus, there may be more subtleties to the association of DIV-1 infection of Daphnia with ecological and evolutionary consequences, such as costs to resistance or covert infection, that the authors acknowledge, and that would be benefitted by coupling experimental and observational studies with the determination of actual infection and viral loads.
References
Prosnier L., N. Loeuille, F.D. Hulot, D. Renault, C. Piscart, B. Bicocchi, M, Deparis, M. Lam, & V. Médoc. (2023). Parasites make hosts more profitable but less available to predators. BioRxiv, ver. 4 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2022.02.08.479552
Recommendations to address uncertainties in environmental risk assessment using toxicokinetics-toxicodynamics models
Addressing uncertainty in Environmental Risk Assessment using mechanistic toxicological models coupled with Bayesian inference
Recommended by Luis Schiesari based on reviews by Andreas Focks and 2 anonymous reviewersEnvironmental Risk Assessment (ERA) is a strategic conceptual framework to characterize the nature and magnitude of risks, to humans and biodiversity, of the release of chemical contaminants in the environment. Several measures have been suggested to enhance the science and application of ERA, including the identification and acknowledgment of uncertainties that potentially influence the outcome of risk assessments, and the appropriate consideration of temporal scale and its linkage to assessment endpoints [1].
Baudrot & Charles [2] proposed to approach these questions by coupling toxicokinetics-toxicodynamics models, which describe the time-course of processes leading to the adverse effects of a toxicant, with Bayesian inference. TKTD models separate processes influencing an organismal internal exposure (´toxicokinetics´, i.e., the uptake, bioaccumulation, distribution, biotransformation and elimination of a toxicant) from processes leading to adverse effects and ultimately its death (´toxicodynamics´) [3]. Although species and substance specific, the mechanistic nature of TKTD models facilitates the comparison of different toxicants, species, life stages, environmental conditions and endpoints [4].
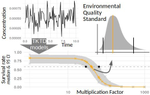
Baudrot & Charles [2] investigated the use of a Bayesian framework to assess the uncertainties surrounding the calibration of General Unified Threshold Models of Survival (a category of TKTD) with data from standard toxicity tests, and their propagation to predictions of regulatory toxicity endpoints such as LC(x,t) [the lethal concentration affecting any x% of the population at any given exposure duration of time t] and MF(x,t) [an exposure multiplication factor leading to any x% effect reduction due to the contaminant at any time t].
Once calibrated with empirical data, GUTS models were used to explore individual survival over time, and under untested exposure conditions. Lethal concentrations displayed a strong curvilinear decline with time of exposure. For a given total amount of contaminant, pulses separated by short time intervals yielded higher mortality than pulses separated by long time intervals, as did few pulses of high amplitude when compared to multiple pulses of low amplitude. The response to a pulsed contaminant exposure was strongly influenced by contaminant depuration times. These findings highlight one important contribution of TKTD modelling in ecotoxicology: they represent just a few of the hundreds of exposure scenarios that could be mathematically explored, and that would be unfeasible or even unethical to conduct experimentally.
GUTS models were also used for interpolations or extrapolations of assessment endpoints, and their marginal distributions. A case in point is the incipient lethal concentration. The responses of model organisms to contaminants in standard toxicity tests are typically assessed at fixed times of exposure (e.g. 24h or 48h in the Daphnia magna acute toxicity test). However, because lethal concentrations are strongly time-dependent, it has been suggested that a more meaningful endpoint would be the incipient (i.e. asymptotic) lethal concentration when time of exposure increases to infinity. The authors present a mathematical solution for calculating the marginal distribution of such incipient lethal concentration, thereby providing both more relevant information and a way of comparing experiments, compounds or species tested for different periods of time.
Uncertainties were found to change drastically with time of exposure, being maximal at extreme values of x for both LC(x,t) and MF(x,t). In practice this means that assessment endpoints estimated when the effects of the contaminant are weak (such as LC10, the contaminant concentration resulting in the mortality of 10% of the experimental population), a commonly used assessment value in ERA, are prone to be highly variable.
The authors end with recommendations for improved experimental design, including (i) using assessment endpoints at intermediate values of x (e.g., LC50 instead of LC10) (ii) prolonging exposure and recording mortality over the course of the experiment (iii) experimenting one or few peaks of high amplitude close to each other when assessing pulsed exposure. Whereas these recommendations are not that different from current practices, they are based on a more coherent mechanistic grounding.
Overall, this and other contributions from Charles, Baudrot and their research group contribute to turn TKTD models into a real tool for Environmental Risk Assessment. Further enhancement of ERA´s science and application could be achieved by extending the use of TKTD models to sublethal rather than lethal effects, and to chronic rather than acute exposure, as these are more controversial issues in decision-making regarding contaminated sites.
References
[1] Dale, V. H., Biddinger, G. R., Newman, M. C., Oris, J. T., Suter, G. W., Thompson, T., ... & Chapman, P. M. (2008). Enhancing the ecological risk assessment process. Integrated environmental assessment and management, 4(3), 306-313. doi: 10.1897/IEAM_2007-066.1
[2] Baudrot, V., & Charles, S. (2018). Recommendations to address uncertainties in environmental risk assessment using toxicokinetics-toxicodynamics models. bioRxiv, 356469, ver. 3 peer-reviewed and recommended by PCI Ecol. doi: 10.1101/356469
[3] EFSA Panel on Plant Protection Products and their Residues (PPR), Ockleford, C., Adriaanse, P., Berny, P., Brock, T., Duquesne, S., ... & Kuhl, T. (2018). Scientific Opinion on the state of the art of Toxicokinetic/Toxicodynamic (TKTD) effect models for regulatory risk assessment of pesticides for aquatic organisms. EFSA Journal, 16(8), e05377. doi: 10.2903/j.efsa.2018.5377
[4] Jager, T., Albert, C., Preuss, T. G., & Ashauer, R. (2011). General unified threshold model of survival-a toxicokinetic-toxicodynamic framework for ecotoxicology. Environmental science & technology, 45(7), 2529-2540. doi: 10.1021/es103092a