
The spatial dynamics of habitat fragmentation drives the evolution of dispersal and metapopulation persistence
Evolution of dispersal and the maintenance of fragmented metapopulations
Abstract
Recommendation: posted 14 July 2023, validated 15 July 2023
Guichard, F. (2023) The spatial dynamics of habitat fragmentation drives the evolution of dispersal and metapopulation persistence. Peer Community in Ecology, 100426. https://doi.org/10.24072/pci.ecology.100426
Recommendation
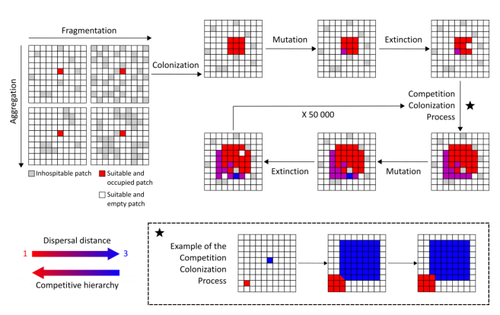
The persistence of populations facing the destruction of their habitat is a multifaceted question that has mobilized theoreticians and empiricists alike for decades. As an ecological question, persistence has been studied as the spatial rescue of populations via dispersal into remaining suitable habitats. The spatial aggregation of habitat destruction has been a key component of these studies, and it has been applied to the problem of coexistence by integrating competition-colonization tradeoffs. There is a rich ecological literature on this topic, both from theoretical and field studies (Fahrig 2003). The relationship between life-history strategies of species and their resilience to spatially structured habitat fragmentation is also an important component of conservation strategies through the management of land use, networks of protected areas, and the creation of corridors. In the context of environmental change, the ability of species to adapt to changes in landscape configuration and availability can be treated as an eco-evolutionary process by considering the possibility of evolutionary rescue (Heino and Hanski 2001; Bell 2017). However, eco-evolutionary dynamics considering spatially structured changes in landscapes and life-history tradeoffs remains an outstanding question. Finand et al. (2023) formulate the problem of persistence in fragmented landscapes over evolutionary time scales by studying models for the evolution of dispersal in relation to habitat fragmentation and spatial aggregation. Their simulations were conducted on a spatial grid where individuals can colonize suitable patch as a function of their competitive rank that decreases as a function of their (ii) dispersal distance trait. Simulations were run under fixed habitat fragmentation (proportion of unsuitable habitat) and aggregation, and with an explicit rate of habitat destruction to study evolutionary rescue.
Their results reveal a balance between the selection for high dispersal under increasing habitat fragmentation and selection for lower dispersal in response to habitat aggregation. This balance leads to the coexistence of polymorphic dispersal strategies in highly aggregated landscapes with low fragmentation where high dispersers inhabit aggregated habitats while low dispersers are found in isolated habitats. The authors then integrate the spatial rescue mechanism to the problem of evolutionary rescue in response to temporally increasing fragmentation. There they show how rapid evolution allows for evolutionary rescue through the evolution of high dispersal. They also show the limits to this evolutionary rescue to cases where both aggregation and fragmentation are not too high. Interestingly, habitat aggregation prevents evolutionary rescue by directly affecting the evolutionary potential of dispersal. The study is based on simple scenarios that ignore the complexity of relationships between dispersal, landscape properties, and species interactions. This simplicity is the strength of the study, revealing basic mechanisms that can now be tested against other life-history tradeoffs and species interactions. Finand et al. (2023) provide a novel foundation for the study of eco-evolutionary dynamics in metacommunities exposed to spatially structured habitat destruction. They point to important assumptions that must be made along the way, including the relationships between dispersal distance and fecundity (they assume a positive relationship), and the nature of life-history tradeoffs between dispersal rate and local competitive abilities.
References
Bell, G. 2017. Evolutionary Rescue. Annual Review of Ecology, Evolution, and Systematics 48:605–627. https://doi.org/10.1146/annurev-ecolsys-110316-023011
Fahrig, L. 2003. Effects of Habitat Fragmentation on Biodiversity. Annual Review of Ecology, Evolution, and Systematics 34:487–515. https://doi.org/10.2307/30033784
Finand, B., T. Monnin, and N. Loeuille. 2023. Evolution of dispersal and the maintenance of fragmented metapopulations. bioRxiv, 2022.06.08.495260, ver. 3 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2022.06.08.495260
Heino, M., and I. Hanski. 2001. Evolution of Migration Rate in a Spatially Realistic Metapopulation Model. The American Naturalist 157:495–511. https://doi.org/10.1086/319927

The recommender in charge of the evaluation of the article and the reviewers declared that they have no conflict of interest (as defined in the code of conduct of PCI) with the authors or with the content of the article. The authors declared that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.
No indication
Evaluation round #2
DOI or URL of the preprint: https://doi.org/10.1101/2022.06.08.495260
Version of the preprint: 2
Author's Reply, 13 Jul 2023
Final suggestions
I am almost ready to make my final recommendation. While I am finalizing it, I simply wanted the authors to consider the following suggestions:
We thank you for your new comments and used them to improve our manuscript. We detail below how we handled them.
1. Competition-colonization trade-off is a central ecological context for the study. Reviewers suggested a number of references that could help set that context. While I understand competition-colonization tradeoffs constitutes the context rather than the goal of the study, the fact that the study focuses on evolutionary process makes it all the more important to provide appropriate reference to the ecological literature that might be unfamiliar to evolutionary biologists.
We have followed this advice in the new version of the manuscript. We have added a sentence in the introduction that provides some of the references proposed in the previous round of reviews. (L78-83).
2. L227-228: vague description of stationarity. Long term average and variance would be more specific and appropriate metrics. In any case, consider providing a more precise and repeatable description of your assessment of stationarity.
We understand that the definition of what we call stationary is not clear. We have added a sentence to highlight that stationarity is that the trait distribution (more precisely its mean and variance) is stable over 5 000 time step at least (218-222). An exact threshold is difficult to apply due to the stochastic components of our model, especially at high levels of fragmentation (hence our discussion on drift). We have added a supplementary figure and a text within the supp info to discuss this. We show different examples of simulations and highlight how our simulation time is sufficient to reach a stable regime (Figure S2).
3. L458. Replace ‘is founded’ with ‘was funded’.
Thanks. Corrected.
Decision by Frédéric Guichard, posted 02 Jun 2023, validated 05 Jun 2023
I am almost ready to make my final recommendation. While I am finalizing it, I simply wanted the authors to consider the following suggestions:
1. Competition-colonization trade-off is a central ecological context for the study. Reviewers suggested a number of references that could help set that context. While I understand competition-colonization tradeoffs constitutes the context rather than the goal of the study, the fact that the study focuses on evolutionary process makes it all the more important to provide appropriate reference to the ecological literature that might be unfamiliar to evolutionary biologists.
2. L227-228: vague description of stationarity. Long term average and variance would be more specific and appropriate metrics. In any case, consider providing a more precise and repeatable description of your assessment of stationarity.
3. L458. Replace ‘is founded’ with ‘was funded’.
Evaluation round #1
DOI or URL of the preprint: https://doi.org/10.1101/2022.06.08.495260
Version of the preprint: 1
Author's Reply, 22 May 2023
Decision by Frédéric Guichard, posted 09 Sep 2022
All reviewers found the work novel and of general interest. I share that opinion. They also provided detailed comments and suggestions to improve the manuscript. Most major concerns converged among reviewers to questions of clarity/reproducilbility, reference to previous work, and interpretation of results. The detailed suggestions found in reviewers' reports will most certainly contribute to improving the manuscript. Reviewers were concerns the work would not be reproducible becasue of lack of details on the various packages and code used to run simulations. Most reviewers also raised concerns over some interpretations of results generated from a specific model. Part of the problem was a concern over the sensitivity of results to the specific model used. Some more specific potential issues were also suggested, such as the dependence of results on the asumed correlation between dispersal and fecundity, or the validity of concluding to lower competition in highly fragmented landscapes. Finally, all reviewers suggested additional references to previous work to better set the broader context for adding evolutionary dynamics to the study of extinctions in metapopulations with habilitat destruction. In my opinion, addressing these, and all other comments from reviewers, will increase the potential impact of this study.
Reviewed by Eva Kisdi , 01 Jul 2022
, 01 Jul 2022
This paper investigates the evolution of dispersal via numerical simulations in a spatially explicit system, assuming that dispersal is traded off with competitive ability. The paper has potential, but some simulations can be criticized for having very few individuals. The interpretation of results and comparison to earlier work should also be improved.
(1) Many results pertain to the case when 99% of the landscape is unsuitable. In this case, there are only 25 patches that can support an individual. How much genetic drift plays a role at such low numbers? How much are the results repeatable over different lattices? with only 25 patches, there is considerable randomness in their actual locations and distances.
(2) The authors say that fragmentation decreases overall occupancy and therefore reduces competition (line 311). Figure 3, however, shows almost 100% occupancy of habitable patches even at a very high level of fragmentation.
(3) The comparison to earlier results in lines 295-309 neglects the fact that in the present model, higher dispersal also means higher fecundity: A dispersive strategy can colonize many patches in the absence of the competitively superior strategy. Many previous models assumed, however, that fecundity is constant and dispersal affects only the spatial allocation of offspring. Environmental heterogeneity selects against dispersal in these models because dispersing offspring end up, on average, in a worse place than where they come from. Dispersal would not be selected against in these previous models if the dispersing offspring were extra offspring in addition to those who do not disperse. In the present model, this latter situation is the case and dispersal would be highly favoured also without the competition-colonization trade-off. (The competition-colonization trade-off explains in the present model why low dispersal may be present.)
I believe the same difference explains why fragmentation decreases dispersal in many natural systems (lines 321-323). If a parent has only so many offspring, then wasting them to disperse into mostly uninhabitable patches is selected against. Instead, the authors' model focuses on a highly fecund organism, which can afford to disperse some of its offspring even if most dispersed offspring perish in unsuitable patches. This is because most of the non-dispersed offspring would perish to kin competition anyway (Hamilton and May 1977); it does not matter how many offspring remain in the patch, the patch will be inhabited by one surviving individual, i.e., it's siblings will die.
The same issue arises also when comparing dispersal dimorphism to Bonte et al (2010): they assumed that the total number of offspring is independent of the dispersal strategy, the present model assumes otherwise. The other papers cited here consider the probability of dispersal with a global dispersal pool, not dispersal distance on a lattice. The comparison should therefore be done more carefully.
(4) The two reasons given in lines 367-372 are in fact the same: with sufficiently strong aggregation, like the cases described in line 383-392, fragmentation does not affect the population and therefore neither drives it extinct nor selects for different dispersal. Strongly aggregated habitat removal is like comparing two lattices of different sizes but both large – they give the same results.
(5) The paper would much benefit from restructuring. The ms investigates evolution in a temporally constant environment and evolutionary rescue in a deteriorating environment. It would help if these were two main sections and the reader would not need to jump back and forth between the two scenarios. The results and their interpretations should be given in one place (currently all interpretation is left to the Discussion). It is a lot easier to grasp and remember results when they make sense! that is, when they are properly explained at the same place where they are described. For one example, the Results section only states the fact that no evolutionary rescue is seen in aggregated landscapes. The explanation is easy; but the reader learns it only almost a hundred lines later, in the second half of the Discussion.
Other comments
- I wonder how much the evolution of dispersal strategies with distance >1 depends on whether the landscape percolates (this may be a highly nontrivial question).
- The authors say that evolutionary rescue is a threshold phenomenon. This is not entirely true, even if there is a jump in Figure 4.
- In theory papers, a section "Materials and methods" really covers the description of the model, so it is better to give it a more suitable title.
- Lines 174-186 repeat what has already been said.
- The legend of figure 2 seems to be mistaken, panels c and d are not what the legend says.
- Panels a and d of figure 3 are not informative.
- Note a typo on the horizontal axis in Figure 4, the last number is presumably 0.1, not 0.01.
- The writing could be improved at some places. E.g. "variations of various selective pressures" does not read well. In line 87, "Fragmentation" is confusing as it may refer to the splitting of colonies described in the previous sentence (replace with habitat fragmentation or start new paragraph). In line 313, " fragmentation intensifies the strength of the competition for space" is confusing becuse the previous sentence said fragmentation decreases competition. The plural of "offspring" is also "offspring"; "evolution" has no plural.
https://doi.org/10.24072/pci.ecology.100426.rev11Reviewed by David Murray-Stoker, 24 Jul 2022
Reviewed by anonymous reviewer 1, 23 Aug 2022
Dear authors,
I have carefully read the paper. The paper deals with the evolution of dispersal in the presence of a colonization-competion tradeoff in the context of a spatially explicit model. The rationale of it is very clear, but model specification is poor. I point to some aspects of the model that I think they are poorly described:
1. How is the definition of the colonization-competition tradeoff set up? In Tilman (1994), the tradeoff is chosen in a very particular way for the whole commuity of S species to coexist.
2. Is the dynamics time discrete?
3. How many possible different dispersal strategies are considered?
4. How the quasi stationary state is checked for? In other words, why 50000 time steps? Aren't the length of the transients depending on parameter values?
In principle, I like the verbal descriptin found in the ms, but a more detailed description would be needed. In any case, what the ms currently lacks is an argument defending that the details of model specification are not relevat for the reported results. In other words, their results are robust to slight changes in model definiton.
In the introduction section, a better connection with previous literature should be achieved. What gives credit to a simulation model is its connections (in some reasonable limits) to existing models. The approach the authors take stems from a stochastic version of Levins metapolation model with habitat destruction. Previous models of this kind have been analyzed in the literature:
Otso Ovaskainen, Kazunori Sato, Jordi Bascompte, and Ilkka Hanski (2002). Metapopulation models for extinction threshold in spatially correlated landscapes. Journal of theoretical Biology, 215: 95-108.
Jordi Bascompte and Miguel A. Rodríguez (2001). Habitat patchiness and plant species richness. Ecology Letters, 4: 417-420.
Jordi Bascompte and Ricard V. Solé (1996). Habitat fragmentation and extinction thresholds in spatially explicit models. Journal of Animal Ecology, 65: 465-473.
Alonso, D., & Mckane, A. (2002). Extinction dynamics in mainland-island metapopulations: an N-patch stochastic model. Bulletin of Mathematical Biology, 64(5), 913–958.
https://doi.org/10.1006/bulm.2002.0307
Solé, R. V., Alonso, D., & Saldaña, J. (2004). Habitat fragmentation and biodiversity collapse in neutral communities. Ecological Complexity, 1(1), 65–75. https://doi.org/10.1016/j.ecocom.2003.12.003
Allouche, O., & Kadmon, R. (2009). A general framework for neutral models of community dynamics. Ecology Letters, 12(12), 1287–1297. https://doi.org/10.1111/j.1461-0248.2009.01379.x
Some of these papers should be cited, because also deal with extinction thresholds in metapopulation models.
I would advice that the authors should start with a simple mean-field one-species model able to recover model results when destruction/recovery and colonization/extinction processes are at random. This would connect to Levins model. From here, introduce S equivalent species, which would correspond to the S-species open Levins model analyzed by Allouche and Kadmon (2009) or Sole etal (2004). Then, introduce S different strategies according to a given colonization-extinction tradeoff, still in the contex of ODE-based patch occupancy metacommunity models, which should correspond to Tilman (1994) with habitat destruction. Then, continue by the introduction of explicit space (a system of N sites or patches), that can be in three states, either destroyed (D), or free but suitable for colonization (A), and fully colonized by a species that belong to any of the S stragies (S_i). And finally, introduce the possibility of evolution.
This close relation of the simulation model presented in the ms to previous literature should be emphasized. It is what gives credit to the model. In some of these limits, do your simulations match model predictions? If yes, you are in a good shape.
Otherwise, in principle, the reader (or this reviewer) should be able to check every line of code to see to what extend results hold or depend on the details of model specification.
BTW, in the context of open science, the model code should be made availabe to check and reuse, by giving credit to the authors, of course.
https://doi.org/10.24072/pci.ecology.100426.rev13
Reviewed by Shripad Tuljapurkar, 05 Sep 2022
Review of Evolution of dispersal and the maintenance of fragmented metapopulations
by: Basile Finand et al.
This paper starts by pointing out that the "mean-field" approach to landscapes, where one considers only the fraction of "suitable" patches, is limited. What also matters is where patches are relative to each other -- i.e., aggregation. The paper then looks at these two dimensions, here called fragmentation and aggregation, as they effect the competition/colonization trade-off first shown by Tilman.
The main phenotype considered is "dispersal" ability, and the authors then also consider phenotypic "evolution" by assuming that random mutations cause a random step change in dispersal distance.
The main results are that fragmentation and aggregation have distinct effects on the competition/colonization trade-off (as described here). In addition, evolution can change dispersal phenotypes fast enough to affect a "rescue" from increasing fragmentation.
Here are my comments.
1. Considerable effort has been taken with the writing so that technical and even biological details are summarized as simply as possible, as is the actual simulation procedure. This is good, but more specifics may be needed, and some of my questions are driven by that.
2. Line 49-50 "fragmentation is defined by the proportion of hostile locations (patches) and we systematically vary its degree of spatial aggregation..."
How about fragmentation in the sense of distance between suitable patches? Is that subsumed in the definition here?
3. I am confused by the term "aggregation," The pictures here do provide an intuitive sense of what "aggregation" may be -- but the reference to R packages does not help.
How about some (simple) statistics, say the spatial correlation, or the conditional probability of finding a similar patch as a function of distance?
4. Does the "scale" of aggregation/fragmentation matter? And how does that connect to the scale of the "patch", if a patch only supports one individual?
5. Line 73 - 78 suggests that Dispersal = Colonization ability? True?
Line 190 says, "competitive hierarchy favors the strategy that has the smallest dispersal distance." Is competition also determined by dispersal?
Presumably then one can say that the results trade-off increased dispersal (good as fragmentation increases) vs. decreased dispersal (competition; good as aggregation increases).
6. Line 262 -- "random few suitable patches" makes no sense.
7. The cited paper by Sciani et al. talks about the R package. While that is good, more detail and discussion (in terms of the underlying theory of how one represents and models landscapes, say the work by Gardner) is essential. Remember that simulations are not proofs, especially simulations that rely on a poorly described process (FBMs, fractals).
https://doi.org/10.24072/pci.ecology.100426.rev14



