
PAQUET Matthieu
- SETE, CNRS, Moulis, France
- Behaviour & Ethology, Demography, Evolutionary ecology, Life history, Maternal effects, Phenotypic plasticity, Population ecology, Statistical ecology
- recommender
Recommendations: 6
Review: 1
Recommendations: 6
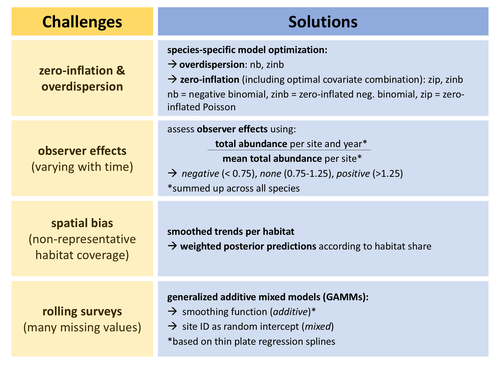
Bird population trend analyses for a monitoring scheme with a highly structured sampling design
Discarding data or dealing with bias?
Recommended by Matthieu Paquet based on reviews by 2 anonymous reviewersObtaining accurate estimates of population trends is crucial to assess populations’ status and make more informed decisions, notably for conservation measures. However, analyzing data we have at hand, including data from systematic monitoring programs, typically induces some bias one way or another (Buckland and Johnston 2017). For example, sampling can be biased towards some types of environments (sometimes historically, before being realized and corrected), and observer identity and experience can vary through time (e.g., an increase in observed experience, if ignored, would cause bias towards positive trends). One way to deal with such biases can be to discard some data, for example, from some overrepresented habitats or from first years surveys to minimize observer bias. However, this may lead to sample sizes becoming too small to detect any trends of interest, especially for surveys with already small temporal resolution (e.g., if time series are too short or with too many missing years).
In this study, Rieger et al. (2025) analyzed data from bird surveys from the Ecological Area Sampling in the German federal state North Rhine-Westphalia in order to assess population trends. This survey uses a ‘rolling’ design, meaning that each site is only visited one year within a multi-year rotation (here six), but this allows to cover a high number of sites. To deal with spatial bias, they analyzed trends per natural region. To control for observer effects, they used a correction factor as an explanatory variable (based on the ratio between the total abundance of all species per site per survey year and the mean total abundance on the same site across all survey years). To deal with the fact that count data for some species but not others may be zero inflated and/or over dispersed, they performed species-specific optimization regarding data distribution (and also regarding inclusion of continuous and categorical covariates). Finally, they deal with the many missing values per year per site (due to the rolling design) by using generalized additive mixed models with site identity as a random intercept.
Importantly, the authors assess how accounting for these biases affects estimates (quite strongly so for some species) and study the consistency of the results with trends estimated from the German Common Bird Monitoring scheme using the software TRIM (Pannekoek and van Strien 2001).
I appreciated their cautious interpretation of their results and of the generalizability of their approach to other datasets. I also recommend that the readers read the review history of the preprint (and I take the opportunity to thank the reviewers and the authors again for the very constructive exchange).
References
Buckland, S., and A. Johnston. 2017. Monitoring the biodiversity of regions: Key principles and possible pitfalls. Biological Conservation 214: 23-34. https://doi.org/10.1016/j.biocon.2017.07.034
Pannekoek, J., van Strienand, A. J. 2001. TRIM 3 manual (Trends & Indices for Monitoring Data). CBS Statistics Netherlands, Voorburg, The Netherlands.
Rieger, M. R., Grüneberg, C., Oberhaus, M., Trautmann, S., Parepa, M., Anthes, N., 2025. Bird population trend analyses for a monitoring scheme with a highly structured sampling design. BioRxiv, ver.3 peer-reviewed and recommended by PCI Ecology https://doi.org/10.1101/2024.06.30.601382
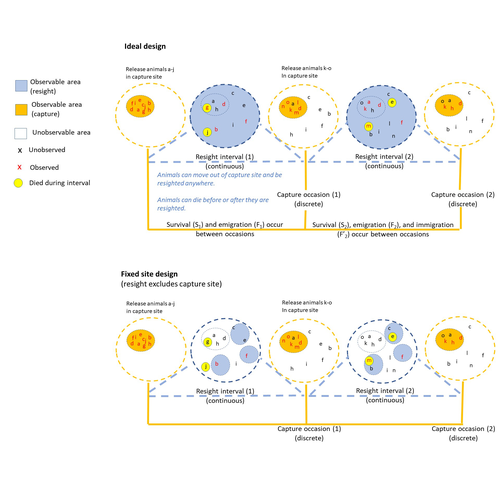
The importance of sampling design for unbiased estimation of survival using joint live-recapture and live resight models
In the quest for estimating true survival
Recommended by Matthieu Paquet based on reviews by Rémi Fay and 1 anonymous reviewerAccurately estimating survival rate and identifying the drivers of its variation is essential for our understanding of population dynamics and life history strategies (Sæther and Bakke 2000), as well as for population management and conservation (Francis et al. 1998, Doherty et al. 2014). Many studies estimate survival from capture–recapture data using the Cormack–Jolly–Seber (CJS) model (Lebreton et al. 1992). However, survival estimates are confounded with permanent emigration from the study area, which can be particularly problematic for mobile species. This is problematic, not only because CJS models under estimate true survival in populations where permanent emigration occurs (i.e. they estimate “apparent” survival), but also because some factors of interest may affect both survival and emigration (e.g., habitat quality, Paquet et al. 2020), leaving the interpretation of results challenging, for example in terms of management decisions.
Several methods have been developed to account for permanent emigration when estimating survival, for example by jointly analyzing CMR data with data on individuals’ locations at each capture/resighting site (to estimate their dispersal distances; Schaub and Royle 2013, Badia Boher et al. 2023), with telemetry data (Powel et al. 2000), mark recovery data (Burnham 1993, Fay et al. 2019), or with live-resight data (Barker 1997).
The Barker joint live-recapture/live-resight (JLRLR) model can estimate survival when resight data are continuous over a long interval and from a larger area than the capture recapture data. This model becomes particularly promising with the growing collection of data from citizen science, or remote detection tools (Dzul et al. 2023). However, as pointed out by Dzul et al., this model assumes that resight probability is homogeneous across the area where individuals can move, and this assumption is likely violated for example because of non-random movements or because of non-random location of resighting sites.
In their manuscript, Dzul et al. performed a thorough simulation study to evaluate the accuracy of survival estimates from JLRLR models under various study designs regarding the location of resight sites (global, random, fixed including the capture site, and fixed excluding the capture site). They simulated data with varying survival and movement values, varying recapture and resight probabilities, and varying sample sizes. Finally, they also developed and fitted a multi state version of the JLRLR model. They show that JLRLR models performed better than CJS models. Survival estimates were still often biased (either positively or negatively) but they were less biased when sesight sites were randomly located (rather than at fixed locations), when recapture sites were included in the resighting design, and when using the multi state JLRLR model they developed.
This study highlights (multistate) JLRLR models as an alternative to CJS models one should consider when auxiliary resight data can be collected. Moreover, it shows the importance of evaluating not only model performance, but also the efficiency of alternative sampling designs before choosing one for our studies. Hopefully, this study will help the authors and other researchers making a more informed and efficient choice of model and design to estimate survival in their study populations.
References
Jaume A. Badia-Boher, Joan Real, Joan Lluís Riera, Frederic Bartumeus, Francesc Parés, Josep Maria Bas, and Antonio Hernández-Matías. Joint estimation of survival and dispersal effectively corrects the permanent emigration bias in mark-recapture analyses. (2023) Scientific reports 13, no. 1: 6970. https://doi.org/10.1038/s41598-023-32866-0
Richard J Barker (1997) Joint modeling of live-recapture, tag-resight, and tag-recovery data. Biometrics: 666-677. https://doi.org/10.2307/2533966
Kenneth P. Burnham (1993) Marked Individuals in the Study of Bird Populations (ed. J.D. Lebreton), pp. 199–213. Birkhäuser, Basel
Kevin E. Doherty, David E. Naugle, Jason D. Tack, Brett L. Walker, Jon M. Graham, Jeffrey L. Beck (2014) Linking conservation actions to demography: grass height explains variation in greater sage‐grouse nest survival. Wildlife biology 20, no. 6 : 320-325. https://doi.org/10.2981/wlb.00004
Maria C. Dzul, Charles B. Yackulic, William L. Kendall (2023) The importance of sampling design for unbiased estimation of survival using joint live-recapture and live resight models. arXiv, ver.3 peer-reviewed and recommended by PCI Ecology https://doi.org/10.48550/arXiv.2312.13414
Rémi Fay, Stephanie Michler, Jacques Laesser, and Michael Schaub (2019) Integrated population model reveals that kestrels breeding in nest boxes operate as a source population. Ecography 42, no. 12: 2122-2131. https://doi.org/10.1111/ecog.04559
Charles M. Francis, John R. Sauer, Jerome R. Serie (1998) Effect of restrictive harvest regulations on survival and recovery rates of American black ducks. The Journal of Wildlife Management : 1544-1557. https://doi.org/10.2307/3802021
Jean-Dominique Lebreton, Kenneth P. Burnham, Jean Clobert, David R. Anderson (1992) Modeling survival and testing biological hypotheses using marked animals: a unified approach with case studies. Ecological monographs 62.1: 67-118. https://doi.org/10.2307/2937171
Matthieu Paquet, Debora Arlt, Jonas Knape, Matthew Low, Pär Forslund, and Tomas Pärt (2020) Why we should care about movements: Using spatially explicit integrated population models to assess habitat source–sink dynamics. Journal of Animal Ecology 89, no. 12: 2922-2933. https://doi.org/10.1111/1365-2656.13357
Larkin A. Powell, Michael J. Conroy, James E. Hines, James D. Nichols, and David G. Krementz. Simultaneous use of mark-recapture and radiotelemetry to estimate survival, movement, and capture rates. (2000) The Journal of Wildlife Management : 302-313. https://doi.org/10.2307/3803003
Bernt-Erik Sæther, Øyvind Bakke (2000) Avian life history variation and contribution of demographic traits to the population growth rate. Ecology 81.3 : 642-653. https://doi.org/10.1890/0012-9658(2000)081[0642:ALHVAC]2.0.CO;2
Michael Schaub, J. Andrew Royle. Estimating true instead of apparent survival using spatial Cormack–Jolly–Seber models (2014) Methods in Ecology and Evolution 5, no. 12: 1316-1326. https://doi.org/10.1111/2041-210X.12134

Sex differences in the relationship between maternal and neonate cortisol in a free-ranging large mammal
Stress and stress hormones’ transmission from mothers to offspring
Recommended by Matthieu Paquet based on reviews by 2 anonymous reviewersIndividuals can respond to environmental changes that they undergo directly (within-generation plasticity) but also through transgenerational plasticity, providing lasting effects that are transmitted to the next generations (Donelson et al. 2012; Munday et al. 2013; Kuijper & Hoyle 2015; Auge et al. 2017, Tariel et al. 2020). These parental effects can affect offspring via various mechanisms, notably via maternal transmission of hormones to the eggs or growing embryos (Mousseau & Fox 1998). While the effects of environmental quality may simply carry-over to the next generation (e.g., females in stressful environments give birth to offspring in poorer condition), parental effects may also be a mechanism that adjusts offspring phenotype in response to environmental variation and predictability, and thereby match offspring's phenotype to future environmental conditions (Gluckman et al. 2005; Marshall & Uller 2007; Dey et al. 2016; Yin et al. 2019), for example by preparing their offspring to an expected stressful environment.
When females experience stress during gestation or egg formation, elevations in glucocorticoids (GC) are expected to affect offspring phenotype in many ways, from the offspring's own GC levels, to their growth and survival (Sheriff et al. 2017). This is a well established idea, but how strong is the evidence for this? A meta-analysis on birds found no clear effect of corticosterone manipulation on offspring traits (38 studies on 9 bird species for corticosterone manipulation; Podmokła et al. 2018). Another meta-analysis including 14 vertebrate species found no clear effect of prenatal stress on offspring GC (Thayer et al. 2018). Finally, a meta-analysis on wild vertebrates (23 species) found no clear effect of GC-mediated maternal effects on offspring traits (MacLeod et al. 2021). As often when facing such inconclusive results, context dependence has been suggested as one potential reason for such inconsistencies, for exemple sex specific effects (Groothuis et al. 2019, 2020). However, sex specific measures on offspring are scarce (Podmokła et al. 2018). Moreover, the literature available is still limited to a few, mostly “model” species.
With their study, Amin et al. (2024) show the way to improve our understanding on GC transmission from mother to offspring and its effects in several aspects. First they used innovative non-invasive methods (which could broaden the range of species available to study) by quantifying cortisol metabolites from faecal samples collected from pregnant females, as proxy for maternal GC level, and relating it to GC levels from hairs of their neonate offspring. Second they used a free ranging large mammal (taxa from which literature is missing): the fallow deer (Dama dama). Third, they provide sex specific measures of GC levels. And finally but importantly, they are exemplary in their transparency regarding 1) the exploratory nature of their study, 2) their statistical thinking and procedure, and 3) the study limitations (e.g., low sample size and high within individual variation of measurements). I hope this study will motivate more research (on the fallow deer, and on other species) to broaden and strengthen our understanding of sex specific effects of maternal stress and CG levels on offspring phenotype and fitness.
References
Amin, B., Fishman, R., Quinn, M., Matas, D., Palme, R., Koren, L., & Ciuti, S. (2024). Sex differences in the relationship between maternal and foetal glucocorticoids in a free-ranging large mammal. bioRxiv, ver. 4 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2023.05.04.538920
Auge, G.A., Leverett, L.D., Edwards, B.R. & Donohue, K. (2017). Adjusting phenotypes via within-and across-generational plasticity. New Phytologist, 216, 343–349. https://doi.org/10.1111/nph.14495
Dey, S., Proulx, S.R. & Teotonio, H. (2016). Adaptation to temporally fluctuating environments by the evolution of maternal effects. PLoS biology, 14, e1002388. https://doi.org/10.1371/journal.pbio.1002388
Donelson, J.M., Munday, P.L., McCormick, M.I. & Pitcher, C.R. (2012). Rapid transgenerational acclimation of a tropical reef fish to climate change. Nature Climate Change, 2, 30. https://doi.org/10.1038/nclimate1323
Gluckman, P.D., Hanson, M.A. & Spencer, H.G. (2005). Predictive adaptive responses and human evolution. Trends in ecology & evolution, 20, 527–533. https://doi.org/10.1016/j.tree.2005.08.001
Groothuis, Ton GG, Bin-Yan Hsu, Neeraj Kumar, and Barbara Tschirren. "Revisiting mechanisms and functions of prenatal hormone-mediated maternal effects using avian species as a model." Philosophical Transactions of the Royal Society B 374, no. 1770 (2019): 20180115. https://doi.org/10.1098/rstb.2018.0115
Groothuis, Ton GG, Neeraj Kumar, and Bin-Yan Hsu. "Explaining discrepancies in the study of maternal effects: the role of context and embryo." Current Opinion in Behavioral Sciences 36 (2020): 185-192. https://doi.org/10.1016/j.cobeha.2020.10.006
Kuijper, B. & Hoyle, R.B. (2015). When to rely on maternal effects and when on phenotypic plasticity? Evolution, 69, 950–968. https://doi.org/10.1111/evo.12635
MacLeod, Kirsty J., Geoffrey M. While, and Tobias Uller. "Viviparous mothers impose stronger glucocorticoid‐mediated maternal stress effects on their offspring than oviparous mothers." Ecology and Evolution 11, no. 23 (2021): 17238-17259.
Marshall, D.J. & Uller, T. (2007). When is a maternal effect adaptive? Oikos, 116, 1957–1963. https://doi.org/10.1111/j.2007.0030-1299.16203.x
Mousseau, T.A. & Fox, C.W. (1998). Maternal effects as adaptations. Oxford University Press.
Munday, P.L., Warner, R.R., Monro, K., Pandolfi, J.M. & Marshall, D.J. (2013). Predicting evolutionary responses to climate change in the sea. Ecology Letters, 16, 1488–1500. https://doi.org/10.1111/ele.12185
Podmokła, Edyta, Szymon M. Drobniak, and Joanna Rutkowska. "Chicken or egg? Outcomes of experimental manipulations of maternally transmitted hormones depend on administration method–a meta‐analysis." Biological Reviews 93, no. 3 (2018): 1499-1517. https://doi.org/10.1111/brv.12406
Sheriff, M. J., Bell, A., Boonstra, R., Dantzer, B., Lavergne, S. G., McGhee, K. E., MacLeod, K. J., Winandy, L., Zimmer, C., & Love, O. P. (2017). Integrating ecological and evolutionary context in the study of maternal stress. Integrative and Comparative Biology, 57(3), 437–449. https://doi.org/10.1093/icb/icx105
Tariel, Juliette, Sandrine Plénet, and Émilien Luquet. "Transgenerational plasticity in the context of predator-prey interactions." Frontiers in Ecology and Evolution 8 (2020): 548660. https://doi.org/10.3389/fevo.2020.548660
Thayer, Zaneta M., Meredith A. Wilson, Andrew W. Kim, and Adrian V. Jaeggi. "Impact of prenatal stress on offspring glucocorticoid levels: A phylogenetic meta-analysis across 14 vertebrate species." Scientific Reports 8, no. 1 (2018): 4942. https://doi.org/10.1038/s41598-018-23169-w
Yin, J., Zhou, M., Lin, Z., Li, Q.Q. & Zhang, Y.-Y. (2019). Transgenerational effects benefit offspring across diverse environments: a meta-analysis in plants and animals. Ecology letters, 22, 1976–1986. https://doi.org/10.1111/ele.13373

The effect of dominance rank on female reproductive success in social mammals
When do dominant females have higher breeding success than subordinates? A meta-analysis across social mammals.
Recommended by Matthieu Paquet based on reviews by 2 anonymous reviewersIn this meta-analysis, Shivani et al. [1] investigate 1) whether dominance and reproductive success are generally associated across social mammals and 2) whether this relationship varies according to a) life history traits (e.g., stronger for species with large litter size), b) ecological conditions (e.g., stronger when resources are limited) and c) the social environment (e.g., stronger for cooperative breeders than for plural breeders). Generally, the results are consistent with their predictions, except there was no clear support for this relationship to be conditional on the ecological conditions. considered
As I have previously recommended the preregistration of this study [2,3], I do not have much to add here, as such recommendation should not depend on the outcome of the study. What I would like to recommend is the whole scientific process performed by the authors, from preregistration sent for peer review, to preprint submission and post-study peer review. It is particularly recommendable to notice that this project was a Masters student project, which shows that it is possible and worthy to preregister studies, even for such rather short-term projects. I strongly congratulate the authors for choosing this process even for an early career short-term project. I think it should be made possible for short-term students to conduct a preregistration study as a research project, without having to present post-study results. I hope this study can encourage a shift in the way we sometimes evaluate students’ projects.
I also recommend the readers to look into the whole pre- and post- study reviewing history of this manuscript and the associated preregistration, as it provides a better understanding of the process and a good example of the associated challenges and benefits [4]. It was a really enriching experience and I encourage others to submit and review preregistrations and registered reports!
References
[1] Shivani, Huchard, E., Lukas, D. (2022). The effect of dominance rank on female reproductive success in social mammals. EcoEvoRxiv, rc8na, ver. 10 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.32942/osf.io/rc8na
[2] Shivani, Huchard, E., Lukas, D. (2020). Preregistration - The effect of dominance rank on female reproductive success in social mammals In principle acceptance by PCI Ecology of the version 1.2 on 07 July 2020. https://dieterlukas.github.io/Preregistration_MetaAnalysis_RankSuccess.html
[3] Paquet, M. (2020) Why are dominant females not always showing higher reproductive success? A preregistration of a meta-analysis on social mammals. Peer Community in Ecology, 100056. https://doi.org/10.24072/pci.ecology.100056
[4] Parker, T., Fraser, H., & Nakagawa, S. (2019). Making conservation science more reliable with preregistration and registered reports. Conservation Biology, 33(4), 747-750. https://doi.org/10.1111/cobi.13342

Sexual coercion in a natural mandrill population
Rare behaviours can have strong effects: evidence for sexual coercion in mandrills
Recommended by Matthieu Paquet based on reviews by Micaela Szykman Gunther and 1 anonymous reviewerSexual coercion can be defined as the use by a male of force, or threat of force, which increases the chances that a female will mate with him at a time when she is likely to be fertile, and/or decrease the chances that she will mate with other males, at some cost to the female (Smuts & Smuts 1993). It has been evidenced in a wide range of species and may play an important role in the evolution of sexual conflict and social systems. However, identifying sexual coercion in natural systems can be particularly challenging. Notably, while male behaviour may have immediate consequences on mating success (“harassment”), the mating benefits may be delayed in time (“intimidation”), and in such cases, evidencing coercion requires detailed temporal data at the individual level. Moreover, in some species male aggressive behaviours may be subtle or rare and hence hardly observed, yet still have important effects on female mating probability and fitness. Therefore, investigating the occurrence and consequences of sexual coercion in such species is particularly relevant but studying it in a statistically robust way is likely to require a considerable amount of time spent observing individuals.
In this paper, Smit et al. (2022) test three clear predictions of the sexual coercion hypothesis in a natural population of Mandrills, where severe male aggression towards females is rare: (1) male aggression is more likely on sexually receptive females than on females in other reproductive states, (2) receptive females are more likely to be injured and (3) male aggression directed towards females is positively related to subsequent probability of copulation between those dyads. They also tested an alternative hypothesis, the “aggressive male phenotype” under which the correlation between male aggression towards females and subsequent mating could be statistically explained by male overall aggressivity. In agreement with the three predictions of the sexual coercion hypothesis, (1) male aggression was on average 5 times more likely, and (2) injuries twice as likely, to be observed on sexually receptive females than on females in other reproductive states and (3) copulation between males and sexually receptive females was twice more likely to be observed when aggression by this male was observed on the female before sexual receptivity. There was no support for the aggressive male hypothesis.
The reviewers and I were highly positive about this study, notably regarding the way it is written and how the predictions are carefully and clearly stated, tested, interpreted, and discussed.
This study is a good illustration of a case where some behaviours may not be common or obvious yet have strong effects and likely important consequences and thus be clearly worth studying. More generally, it shows once more the importance of detailed long-term studies at the individual level for our understanding of the ecology and evolution of wild populations.
It is also a good illustration of the challenges faced, when comparing the likelihood of contrasting hypotheses means we need to alter sample sizes and/or the likelihood to observe at all some behaviours. For example, observing copulation within minutes after aggression (and therefore, showing statistical support for “harassment”) is inevitably less likely than observing copulations on the longer-term (and therefore showing statistical support for “intimidation”, when of course effort is put into recording such behavioural data on the long-term). Such challenges might partly explain some apparently intriguing results. For example, why are swollen females more aggressed by males if only aggression before the swollen period seems associated with more chances of mating? Here, the authors systematically provide effect sizes (and confidence intervals) and often describe the effects in an intuitive biological way (e.g., “Swollen females were, on average, about five times more likely to become injured”). This clearly helps the reader to not merely compare statistical significances but also the biological strengths of the estimated effects and the uncertainty around them. They also clearly acknowledge limits due to sample size when testing the harassment hypothesis, yet they provide precious information on the probability of observing mating (a rare behaviour) directly after aggression (already a rare behaviour!), that is, 3 times out of 38 aggressions observed between a male and a swollen female. Once again, this highlights how important it is to be able to pursue the enormous effort put so far into closely and continuously monitoring this wild population.
Finally, this study raises exciting new questions, notably regarding to what extent females exhibit “counter-strategies” in response to sexual coercion, notably whether there is still scope for female mate choice under such conditions, and what are the fitness consequences of these dynamic conflicting sexual interactions. No doubt these questions will sooner than later be addressed by the authors, and I am looking forward to reading their upcoming work.
References
Smit N, Baniel A, Roura-Torres B, Amblard-Rambert P, Charpentier MJE, Huchard E (2022) Sexual coercion in a natural mandrill population. bioRxiv, 2022.02.07.479393, ver. 5 peer-reviewed and recommended by Peer Community in Ecology. https://doi.org/10.1101/2022.02.07.479393
Smuts BB, Smuts R w. (1993) Male Aggression and Sexual Coercion of Females in Nonhuman Primates and Other Mammals: Evidence and Theoretical Implications. In: Advances in the Study of Behavior (eds Slater PJB, Rosenblatt JS, Snowdon CT, Milinski M), pp. 1–63. Academic Press. https://doi.org/10.1016/S0065-3454(08)60404-0

Preregistration - The effect of dominance rank on female reproductive success in social mammals
Why are dominant females not always showing higher reproductive success? A preregistration of a meta-analysis on social mammals
Recommended by Matthieu Paquet based on reviews by Bonaventura Majolo and 1 anonymous reviewerIn social species conflicts among group members typically lead to the formation of dominance hierarchies with dominant individuals outcompeting other groups members and, in some extreme cases, suppressing reproduction of subordinates. It has therefore been typically assumed that dominant individuals have a higher breeding success than subordinates. However, previous work on mammals (mostly primates) revealed high variation, with some populations showing no evidence for a link between female dominance reproductive success, and a meta-analysis on primates suggests that the strength of this relationship is stronger for species with a longer lifespan [1]. Therefore, there is now a need to understand 1) whether dominance and reproductive success are generally associated across social mammals (and beyond) and 2) which factors explains the variation in the strength (and possibly direction) of this relationship.
In their preregistration, Shivani et al. [2] plan to perform a meta-analysis on 86 social mammal species to address these two points. More specifically, they will investigate whether the relationship between female dominance and reproductive success vary according to life history traits (e.g. stronger for species with large litter size), ecological conditions (e.g. stronger when resources are limited) and the social environment (e.g. stronger for cooperative breeders than for plural breeders).
The two reviewers and I were particularly positive and enthusiastic about this preregistration and only had minor comments that were nicely addressed by the authors. We found the background well-grounded in the existing literature and that the predictions were therefore clear and well-motivated. The methods were particularly transparent with a nicely annotated R script and the authors even simulated a dataset with the same structure as the actual data in order to make sure that the coding of the data handling and statistical analyses were appropriate (without being tempted to look at model outputs from the true dataset).
Perhaps one limitation to keep in mind once we will have the chance to look at the outcome of this study if that the dataset may not be fully representative of social species with dominance hierarchies. For example, the current dataset contains only one aquatic mammal (Mirounga angustirostris) as far as I can see, which is likely due to a lack of knowledge on such systems. Furthermore, not only mammals exhibit dominance hierarchies and it will be interesting to see if the results of the proposed study hold for other social taxa (and if not, what may explain their differences).
That being said, the proposed study will already offer a much broader overview of the relationship between dominance and reproductive success in animal societies and a better understanding for its variation. The reviewers and I believe it will make an important contribution to the fields of socio-ecology and evolutionary ecology. I therefore strongly recommend this preregistration and we are particularly looking forward to seeing the outcome of this exciting study.
References
[1] Majolo, B., Lehmann, J., de Bortoli Vizioli, A., & Schino, G. (2012). Fitness‐related benefits of dominance in primates. American journal of physical anthropology, 147(4), 652-660. doi: 10.1002/ajpa.22031
[2] Shivani, Huchard, E., Lukas, D. (2020). Preregistration - The effect of dominance rank on female reproductive success in social mammals In principle acceptance by PCI Ecology of the version 1.2 on 07 July 2020. https://github.com/dieterlukas/FemaleDominanceReproductionMetaAnalysis/blob/trunk/PreregistrationMetaAnalysis_RankSuccess.Rmd
Review: 1

Investigating the rare behavior of male parental care in great-tailed grackles
Studying a rare behavior in a polygamous bird: male parental care in great-tailed grackles
Recommended by Marie-Jeanne Holveck based on reviews by Matthieu Paquet and André C FerreiraThe Great-tailed grackle (Quiscalus mexicanus) is a polygamous bird species that is originating from Central America and rapidly expanding its geographic range toward the North, and in which females were long thought to be the sole nest builders and caretakers of the young. In their pre-registration [1], Folsom and collaborators report repeated occurrences of male parental care and develop hypotheses aiming at better understanding the occurrence and the fitness consequences of this very rarely observed male behavior. They propose to assess if male parental care correlates with the circulating levels of several relevant hormones, increases offspring survival, is a local adaptation, and is a mating strategy, in surveying three populations located in Arizona (middle of the geographic range expansion), California (northern edge of the geographic range), and in Central America (core of the range). This study is part of a 5-year bigger project.
Both reviewers and I strongly value Folsom and collaborators’ commitment to program a study, in natural field conditions, of a rare, yet likely evolutionary-important behavior, namely parental care by males of the great-tailed grackle. Yet, we all also recognized that it is a risky endeavor, and as a consequence, we wondered about the authors’ ability to reach a sufficient sample size to statistically test (all) hypotheses and predictions with enough confidence (e.g. risk of type I errors, also known as false positives).
Folsom and collaborators acknowledged these limitations in their pre-registration. (i) They made the exploratory nature of their research work clear to readers. (ii) They adapted their analysis plan in running prior power analyses and in focusing on effect sizes (estimates and confidence intervals). (iii) Last and not least, Folsom and collaborators clearly exposed a priori hypotheses, their associated predictions and alternatives, and ranked the latter based on their plausibility according to knowledge in the current and other study systems. Developing theory about male parental care behavior more generally with regard to a polygamous species that is rapidly expanding its geographic range and that is considered not to provide male parental care is without any doubt an added value to this study.
In summary, while this study will likely be insufficient to fully understand male parental care behavior of great-tailed grackles, it is much needed because it will definitely allow rejecting some predictions (e.g., if this behavior is present in all the studied populations, it would be common across range against expectation; finding only one male providing care to an unrelated offspring would lead to reject the prediction that males only care for their own offspring) and thus it will help laying the foundation of future research directions.
I strongly support the pre-registration system and thank all the contributors for producing a fruitful discussion throughout the review process, which in fine improved the clarity and logic of this pre-registration. Given the positive and encouraging reviews, the detailed and thorough answers to all comments by Folsom and collaborators, and their satisfactory and interesting revisions, I am happy to recommend this pre-registration and I look forward to seeing its outcomes.
References
[1] Folsom MA, MacPherson M, Lukas D, McCune KB, Bergeron L, Bond A, Blackwell A, Rowney C, Logan CJ. 2020. Investigating the rare behavior of male parental care in great-tailed grackles. corinalogan.com/Preregistrations/gmalecare.html In principle acceptance by PCI Ecology of the version on 15 June 2020 corinalogan/grackles/blob/master/Files/Preregistrations/gmalecare.Rmd.