
Direct submissions to PCI Ecology from bioRxiv.org are possible using the B2J service
Latest recommendations
| Id | Title * | Authors * | Abstract * | Picture * | Thematic fields * | Recommender | Reviewers | Submission date | |
|---|---|---|---|---|---|---|---|---|---|
26 Mar 2025
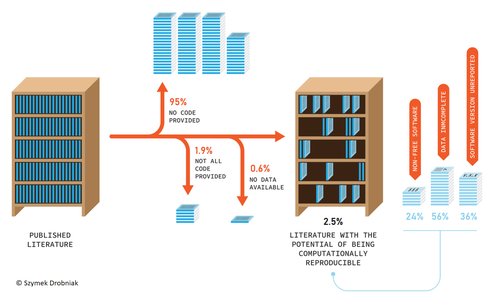
Code-sharing policies are associated with increased reproducibility potential of ecological findingsEnsuring reproducible science requires policiesRecommended by Ignasi Bartomeus based on reviews by Francisco Rodriguez-Sanchez and Veronica Cruz based on reviews by Francisco Rodriguez-Sanchez and Veronica Cruz
Researchers do not live in a vacuum, and the social context we live in affects how we do science. On one hand, increased competition for scarce funding creates the wrong incentives to do fast analysis, leading sometimes to poorly checked results that accumulate errors (Fraser et al. 2018). On the other hand, the actual challenges the world faces require more than ever robust scientific evidence that can be used to tackle the current rapid human-induced environmental change. Moreover, scientists' credibility is at stake at this moment where the global flow of information can be politically manipulated, and accessing reliable sources of information is paramount for society. At the crossroads of these challenges is scientific reproducibility. Making our results transparent and reproducible ensures that no perverse incentives can compromise our findings, that results can be reliably applied to solve relevant problems, and that we regain societal credibility in the scientific process. Unfortunately, in ecology and evolution, we are still far from publishing open, transparent, and reproducible papers (Maitner et al. 2024). Understanding which factors promote increased use of good practices regarding reproducibility is hence very welcome. Sanchez-Tojar and colleagues (2025) conducted a (reproducible) analysis of code and data-sharing practices (a cornerstone of scientific reproducibility) in journals with and without explicit policies regarding data and code deposition. The gist is that having policies in place increases data and code sharing. Doing science about how we do science (meta-science) is important to understand which actions drive our behavior as scientists. This paper highlights that in the absence of strong societal or personal incentives to share code and data, clear policies can catalyze this process. However, in my opinion, policies are a needed first step to consolidate a more permanent change in researchers' behavior regarding reproducible science, but policies alone will not be enough to fix the problem if we do not change also the cultural values around how we publish science. Appealing to inner values, and recognizing science needs to be reproducible to ensure potential errors are easily spotted and corrected requires a deep cultural change. References Fraser, Hannah, Tim Parker, Shinichi Nakagawa, Ashley Barnett, and Fiona Fidler. "Questionable research practices in ecology and evolution." PloS one 13, no. 7 (2018): e0200303. https://doi.org/10.1371/journal.pone.0200303 Alfredo Sánchez-Tójar, Aya Bezine, Marija Purgar, Antica Culina (2025) Code-sharing policies are associated with increased reproducibility potential of ecological findings. EcoEvoRxiv, ver.4 peer-reviewed and recommended by PCI Ecology. https://doi.org/10.32942/X21S7H | Code-sharing policies are associated with increased reproducibility potential of ecological findings | Alfredo Sánchez-Tójar, Aya Bezine, Marija Purgar, Antica Culina | <p>Software code (e.g., analytical code) is increasingly recognized as an important research output because it improves transparency, collaboration, and research credibility. Many scientific journals have introduced code-sharing policies; however,... | | Meta-analyses, Preregistrations, Statistical ecology | Ignasi Bartomeus | 2024-12-11 10:33:13 | ||
20 Mar 2025
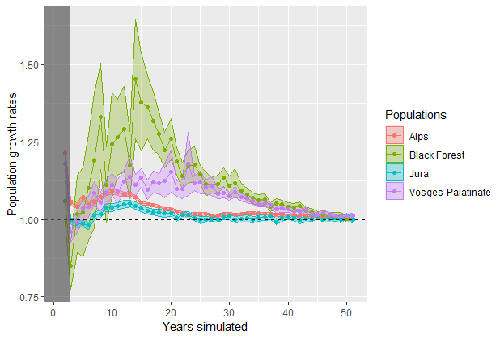
Modelling Eurasian lynx populations in Western Europe: What prospects for the next 50 years?Glimmers of hope for the Eurasian lynx in Western EuropeRecommended by Elodie Vercken based on reviews by Hector Ruiz and Henrik AndrenThe conservation of large carnivores remains a challenge for biodiversity conservation (Ingeman et al. 2022), as they combine strict ecological requirements (large territories, sensitivity to human disturbance) with coexistence conflicts with human activities (livestock farming, hunting, risk perception). Although the Eurasian lynx is currently considered as “least concerned” by the IUCN Red List, this favorable status conceals major disparities between the remaining historical population nuclei in Northern and Eastern Europe and small, isolated populations in Western Europe resulting from reintroduction programs for which long-term persistence remains in jeopardy (Chapron et al. 2014). References Bauduin S, Germain E, Zimmermann F, Idelberger S, Herdtfelder M, Heurich M, Kramer-Schadt S, Duchamp C, Drouet-Hoguet N, Morand A, Blanc L, Charbonnel A, Gimenez O. 2025. Modelling Eurasian lynx populations in Western Europe: What prospects for the next 50 years?https://doi.org/10.1101/2021.10.22.465393 Chapron G, et al. 2014. Recovery of large carnivore in Europe’s modern human-dominated landscapes. Science 345: 1517-1519 https://doi.org/10.1126/science.1257553 Gatti S. 2022. National Action Plan for the Eurasian Lynx: restoring the Lynx to a favorable conservation status un France (2022-2026), 176 p. Ingeman, K.E., Zhao, L.Z., Wolf, C. et al. 2022. Glimmers of hope in large carnivore recoveries.Sci Rep 12, 10005 https://doi.org/10.1038/s41598-022-13671-7 | Modelling Eurasian lynx populations in Western Europe: What prospects for the next 50 years? | Sarah Bauduin, Estelle Germain, Fridolin Zimmermann, Sylvia Idelberger, Micha Herdtfelder, Marco Heurich, Stephanie Kramer-Schadt, Christophe Duchamp, Nolwenn Drouet-Hoguet, Alain Morand, Laetitia Blanc, Anaïs Charbonnel, Olivier Gimenez | <p>Persistence of populations may be uncertain for large carnivore species, especially for those established in human-dominated landscapes. Here, we studied the Eurasian lynx in Western Europe established in the Upper Rhine meta-population (i.e., ... | | Conservation biology, Demography, Dispersal & Migration, Habitat selection, Population ecology, Spatial ecology, Metacommunities & Metapopulations | Elodie Vercken | 2022-01-19 10:17:56 | ||
12 Mar 2025
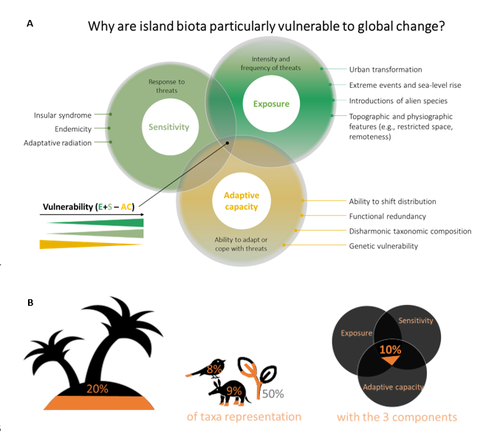
A framework to quantify the vulnerability of insular biota to global changeAssessing vulnerability of island biodiversity to global changeRecommended by Vasilis Dakos based on reviews by T Johnson and 1 anonymous reviewer
Islands may represent just a small fraction (6.67%) of the planet’s land but they host a disproportionate 20% of the world’s biodiversity. Yet islands are highly vulnerable to human-induced change. Out of all IUCN Red list species, almost half of them are found on islands (Russell and Kueffer, 2019) while from the approximately 800 known extinctions that have occurred since the European expansion around the world, 75% have occurred on islands (IUCN, 2017). References Bellard Céline, Marino Clara, Butt Nathalie, Fernández-Palacios José María, Rigal François, Robuchon Marine, Lenoir Jonathan, Irl Severin, Benítez-López Ana, Capdevila Pol, Zhu G, Caetano Gabriel, Denelle Pierre, Philippe-Lesaffre Martin, Schipper Aafke, M Foden Wendy, Kissling W. Daniel, Leclerc Camille (2025) A framework to quantify the vulnerability of insular biota to global change. HAL, ver.3 peer-reviewed and recommended by PCI Ecology https://hal.science/hal-04550966 Frankham, R., Briscoe, D. A., and Ballou, J. D. (2002). Introduction to Conservation Genetics. Cambridge University Press.Fuessel, H.-M. (2007). Vulnerability: A generally applicable conceptual framework for climate change research. Glob. Environ. Change 17, 155–167. https://doi.org/10.1016/j.gloenvcha.2006.05.002 IUCN (2017). IUCN 2017 : International Union for Conservation of Nature annual report 2017. Available at: https://iucn.org/resources/annual-reports/iucn-2017-international-union-conservation-nature-annual-report-2017 (Accessed March 10, 2025). Pacifici, M., Foden, W. B., Visconti, P., Watson, J. E. M., Butchart, S. H. M., Kovacs, K. M., et al. (2015). Assessing species vulnerability to climate change. Nat. Clim. Change 5, 215–224. https://doi.org/10.1038/nclimate2448 Russell, J. C., and Kueffer, C. (2019). Island Biodiversity in the Anthropocene. Annu. Rev. Environ. Resour. 44, 31–60. https://doi.org/10.1146/annurev-environ-101718-033245 | A framework to quantify the vulnerability of insular biota to global change | Bellard Céline, Marino Clara, Butt Nathalie, Fernández-Palacios José María, Rigal François, Robuchon Marine, Lenoir Jonathan, Irl Severin, Benítez-López Ana, Capdevila Pol, Zhu G, Caetano Gabriel, Denelle Pierre, Philippe-Lesaffre Martin, Schipper... | <p>The majority of vulnerability assessments of biodiversity to global changes have so far been applied to, and designed for, mainland systems, overlooking islands. However, islands harbour unique biodiversity and are epicentres of ongoing extinct... | | Biodiversity, Biogeography, Biological invasions, Climate change, Macroecology | Vasilis Dakos | 2024-04-18 15:43:06 | ||
27 Feb 2025
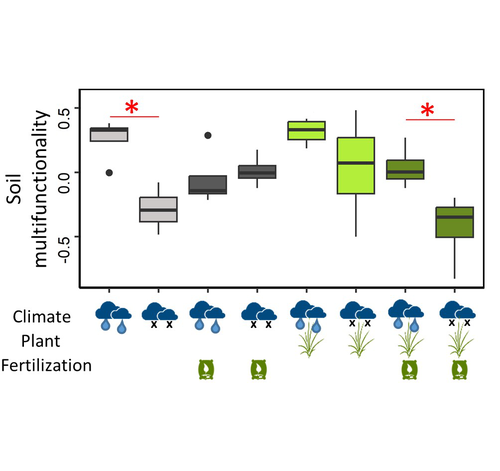
Mineral fertilization reduces the drought resistance of soil multifunctionality in a mountain grassland system through plant-soil interactionsComplex interactions between fertilization, drought and plants impact soil functioningRecommended by Sébastien Barot based on reviews by 2 anonymous reviewers
The ingredients of this study are classic in soil ecology and in the study of belowground-aboveground interactions: the presence of plants, draught and mineral fertilization (for the experimental treatments) and microbial carbon, microbial nitrogen, microbial phosphorus, substrate-induced respiration, cumulative extracellular enzyme activity, nitrogen mineralization potential, nitrification potential, denitrification potential (as a result of the treatments). It is interesting and useful to have tested all the combinations of the three treatments and the height variables (also in the form of a soil multifunctionality index) in the same study and to have been able to express hypotheses on the underlying mechanisms of interaction.
A key result is that mineral fertilization can reduce the soil ability to withstand draughts in terms of soil multifunctionality. This effect would be due to the increase in plant growth associated with fertilization, which reduces the availability of soil resources. This subsequently affects microbial diversity and soil multifunctionality. This confirms that the interactions between plants and soil microorganisms are complex and relevant for understanding and predicting the impact of climate and fertilization on soil functioning and the sustainability of plant-soil systems.
Although the study is rather fundamental, it has been designed to be relevant to grassland management and points to very general mechanisms that are likely to be relevant to arable land management. It would therefore be useful to repeat this work for interactions between a crop and its soil. Finally, it would be crucial to test the impact of heavy fertilization in intensive cropping systems on the resistance and resilience of soil functions to climate variability and climate changes.
A slightly disturbing fact is that the underlying interactions are probably so complicated that it seems so far impossible to me to make predictions about the ranking of the height combinations of treatments on each soil variable. But this complexity is clearly inherent to ecology and, in particular, plant-soil interactions.
References Gabin Piton, Arnaud Foulquier, Lionel Bernard, Aurelie Bonin, Thomas Pommier, Sandra Lavorel, Roberto Geremia, Jean Christophe Clement (2025) Mineral fertilization reduces the drought resistance of soil multifunctionality in a mountain grassland system through plant-soil interactions. bioRxiv, ver.2 peer-reviewed and recommended by PCI Ecology https://doi.org/10.1101/2024.09.19.613911 | Mineral fertilization reduces the drought resistance of soil multifunctionality in a mountain grassland system through plant-soil interactions | Gabin Piton, Arnaud Foulquier, Lionel Bernard, Aurelie Bonin, Thomas Pommier, Sandra Lavorel, Roberto Geremia, Jean Christophe Clement | <p>Increasing droughts threaten soil microbial communities and the multiple functions they control in agricultural soils. These soils are often fertilized with mineral nutrients, but it remains unclear how this fertilization may alter the capacity... | | Agroecology, Climate change, Ecological stoichiometry, Ecosystem functioning, Experimental ecology, Microbial ecology & microbiology, Soil ecology | Sébastien Barot | 2024-09-19 18:55:06 | ||
24 Feb 2025

Drivers of plant-associated invertebrate community structure in West-European coastal dunesCombining Joint Species Distribution Models and multivariate techniques allows understanding biogeographical and micro-habitat community responsesRecommended by Joaquín Hortal based on reviews by Sergio Chozas, André Mira and 1 anonymous reviewer
Community structure is determined by the regional species pool – which for simplicity can be assumed to be filtered through dispersal limitations, abiotic conditions, and species coexistence mechanisms (Cornell & Harrison 2014). This filtering involves macroecological constraints, such as energy and space availability, and assembly rules that determine species composition (Diamond 1975; Weiher & Keddy 1995; Guisan & Rahbek 2011; Hortal et al. 2012). But also by a series of processes that determine species distributions across scales, including biogeographical and stochastic processes (e.g., large-scale dispersal and occupancy dynamics within the landscape) and deterministic niche-based responses to abiotic and biotic conditions, which interact across scales (Soberón 2010; Hortal et al. 2010; Brousseau et al. 2018). These processes collectively determine the persistence of species assemblages within communities. It follows that, to understand the processes determining the structure of these communities it is necessary to combine methods analyse the effects of drivers acting on both species distributions and community responses. Van de Walle et al. (2025) take this integrative approach. The final revised version of their work combines multivariate techniques (in this case a RDA) and Joint SDMs to model the small-scale distribution and structure of the invertebrate communities inhabiting a series of coastal dunes in Southern England, France, Belgium and the Netherlands. The paper builds upon well-designed stratified field surveys, which allow them to identify variations at different scales, from geographical to local. These high-quality field data, together with the combination of different modelling techniques, allows them to identify both a clear biogeographical zonation in the structure of these communities, and the existence of a series of neat responses of species to the spatial structure and vigour of the tussocks created by the marram grass fixing the sand dunes. Their models also include the body size, feeding guild and phylogenetic relationships between co-occurring species, although their effects are smaller compared to those of biogeographical differences –which, arguably, are determined by differences in the species pool of each dune system, and species responses to the microhabitat conditions created by the tussocks. They can however identify a trade-off between generalist and specialist species within each community. Note that here I'm using model in the sense of tools for understanding and explaining complex ecological systems, as advocated by Levins (1966). Which is precisely what Van de Walle et al. (2025) do here. By combining techniques tailored to model species distributions and community-level responses, they (we) gain a much improved understanding of how both species pools and the spatial structure of habitats determine the composition of ecological communities. Importantly, Van de Walle et al. (2025) use this knowledge to obtain key insights about how to manage and restore these endangered habitats, thereby proving the value of this kind of integrative approaches. References Brousseau, P.-M., Gravel, D., & Handa, I. T. (2018). On the development of a predictive functional trait approach for studying terrestrial arthropods. Journal of Animal Ecology, 87(5), 1209–1220. https://doi.org/10.1111/1365-2656.12834 Cornell, H. V., & Harrison, S. P. (2014). What are species pools and when are they important? Annual Review of Ecology, Evolution, and Systematics, 45(1), 45–67. http://dx.doi.org/10.1146/annurev-ecolsys-120213-091759 Diamond, J. M. (1975). Assembly of species communities. In M. L. Cody & J. M. Diamond (Eds.), Ecology and Evolution of Communities (pp. 342–444). Harvard University Press. Guisan, A., & Rahbek, C. (2011). SESAM – a new framework integrating macroecological and species distribution models for predicting spatio-temporal patterns of species assemblages. Journal of Biogeography, 38(8), 1433–1444. https://doi.org/10.1111/j.1365-2699.2011.02550.x Hortal, J., Roura-Pascual, N., Sanders, N. J., & Rahbek, C. (2010). Understanding (insect) species distributions across spatial scales. Ecography, 33(1). https://doi.org/10.1111/j.1600-0587.2009.06428.x Hortal, J., de Marco, P., Santos, A. M. C., & Diniz-Filho, J. A. F. (2012). Integrating biogeographical processes and local community assembly. Journal of Biogeography, 39(4). https://doi.org/10.1111/j.1365-2699.2012.02684.x Levins, R. (1966). The strategy of model building in population biology. American Scientist, 54, 421–431. Soberón, J. (2010). Niche and area of distribution modeling: A population ecology perspective. Ecography, 33(1), 159–167. https://doi.org/10.1111/j.1600-0587.2009.06074.x van de Walle, R., Dahirel, M., Langeraert, W., Benoit, D., Vantieghem, P., Vandegehuchte, M. L., Massol, F., & Bonte, D. (2025). Drivers of plant-associated invertebrate community structure in West-European coastal dunes. BioRxiv, 2024.06.24.600350, ver.3 peer-reviewed and recommended by PCI Ecology https://doi.org/10.1101/2024.06.24.600350 Weiher, E., & Keddy, P. A. (1995). Assembly rules, null models, and trait dispersion: New questions from old patterns. Oikos, 74(1), 159–164. https://doi.org/10.2307/3545686
| Drivers of plant-associated invertebrate community structure in West-European coastal dunes | Ruben Van De Walle, Maxime Dahirel, Ward Langeraert, Dries Benoit, Pieter Vantieghem, Martijn L. Vandegehuchte, François Massol and Dries Bonte | <p>The organisation of species assemblages is affected by environmental factors acting at different spatial scales. To understand the drivers behind the community structure of invertebrates associated with marram grass -the dominant dune-building ... | | Biodiversity, Biogeography, Spatial ecology, Metacommunities & Metapopulations, Species distributions | Joaquín Hortal | 2024-06-28 10:19:36 | ||
07 Feb 2025
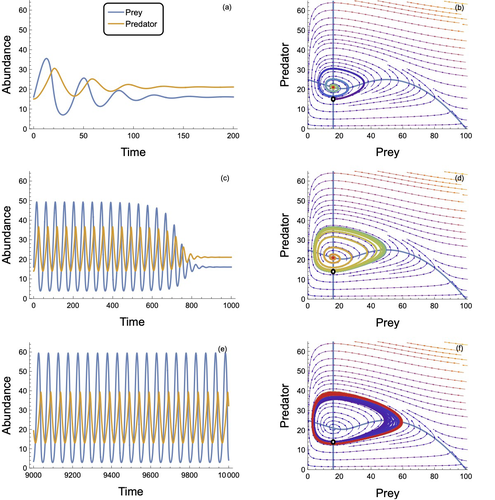
In defense of the original Type I functional response: The frequency and population-dynamic effects of feeding on multiple prey at a timeRevising behavioural assumptions leads to a new appreciation of an old functional response modelRecommended by Frédéric Barraquand based on reviews by Matthieu Barbier and Wojciech Uszko
The functional response, describing the relation between predator intake rate and prey density, is a pivotal concept to understand foraging behaviour and its consequences for community dynamics. Holling (1959a) introduced three types of functional responses according to their shapes, labelled I, II and III. The type II, also known as the disc equation (Holling 1959b), has become popular among empiricists and theoreticians alike, due to its ability to describe predator intake saturation. The type III is often used to represent predator switching to other prey species when main prey density is low. Although theoretical works identify the linear functional response used in Lotka-Volterra models as a type I, Holling (1959a)’s type I model actually envisioned that at some threshold prey density, the linear increase in predator intake with prey density would give way to an upper predator intake limit, so that Holling’s type I has a rectilinear shape, with an angle joining straight lines. Ecology students can actually see this rectilinear shape reproduced in some texbooks, although not in textbook dynamical models, as they usually transition from Lotka-Volterra models to models with type II response. To many, the rectilinear shape of the original type I looks like a historical curiosity: the type II functional response accounts for intake rate saturation with a more convenient smooth function. Novak et al. (2025) turn this preconception on its head by first pedagogically showing that Holling’s original type I model can be obtained as a limit case of a variant of the celebrated type II model. The derivation follows up earlier work by Sjöberg (1980), which might be unfamiliar to readers outside aquatic ecology. The often untold assumption of the type II functional response model is that searching and handling prey are two exclusive behavioural processes, with predators that can only handle one prey item at a time. Allowing for several prey items to be handled at once while searching, until the predator reaches n prey items, the original type I functional response emerges as a limit case of the « multiprey » functional response as n goes to infinity. Interestingly, the multiprey response looks a lot like the original type I for large yet doable n. Novak et al. (2025) then proceed to look for the prevalence of such multiprey functional response shapes in a large database of functional responses (Uiterwaal et al. 2022). Combining linear type I and multiprey models (the asymptote may not always be visible), they find support for this revised type I hypothesis in about one-third of the cases. Although the type II and III models are still well supported by data, the results do suggest that linearity at low prey density may well be more frequent than one thinks. They complement this analysis by showing that larger predators relative to their prey tend to have larger n in the multiprey response. It is consistent with the hypothesis that the bigger you are relative to your prey, the more prey items you can handle at once. Finally, Novak et al. (2025) investigate the consequences of the multiprey model for community dynamics. They find overall a richer dynamical behaviour than the Lotka-Volterra type I and common parameterizations of the type II, suggesting that observed linearity in some range of prey density does not necessarily translate in simpler dynamical behaviour. Novak et al. (2025) provide here a convincing and pedagogical study showing how seemingly benign behavioural assumptions can in fact profoundly alter the perceived relevance of community dynamics models. As they conclude, their analyses have lessons for future empirical functional response work, which should not necessarily dismiss the type I model and consider perhaps variants to the classical type II and III, as well as for future theoretical analyses, which could generalize this model to multiple prey species, or relax other behavioural assumptions. References Holling, C. S. (1959a). The components of predation as revealed by a study of small-mammal predation of the European Pine Sawfly. The Canadian Entomologist, 91(5), 293-320. https://doi.org/10.4039/Ent91293-5 Holling, C. S. (1959b). Some characteristics of simple types of predation and parasitism. The Canadian Entomologist, 91(7), 385-398. https://doi.org/10.4039/Ent91385-7 Novak, M., Coblentz, K. E., & DeLong, J. P (2025). In defense of the original Type I functional response: The frequency and population-dynamic effects of feeding on multiple prey at a time. bioRxiv, ver.4 peer-reviewed and recommended by PCI Ecology https://doi.org/10.1101/2024.05.14.594210 Sjöberg, S. (1980). Zooplankton feeding and queueing theory. Ecological Modelling, 10(3-4), 215-225. https://doi.org/10.1016/0304-3800(80)90060-5 Uiterwaal, S. F., Lagerstrom, I. T., Lyon, S. R., & DeLong, J. P. (2022). FoRAGE database: A compilation of functional responses for consumers and parasitoids. Ecology, 103(7), e3706. https://doi.org/10.1002/ecy.3706 | In defense of the original Type I functional response: The frequency and population-dynamic effects of feeding on multiple prey at a time | Mark Novak, Kyle Edward Coblentz, John P DeLong | <p>Ecologists differ in the degree to which they consider the linear Type I functional response to be an unrealistic versus sufficient representation of predator feeding rates. Empiricists tend to consider it unsuitably non-mechanistic and theoret... | | Coexistence, Community ecology, Food webs, Foraging, Population ecology, Theoretical ecology | Frédéric Barraquand | 2024-05-21 03:44:00 | ||
24 Jan 2025
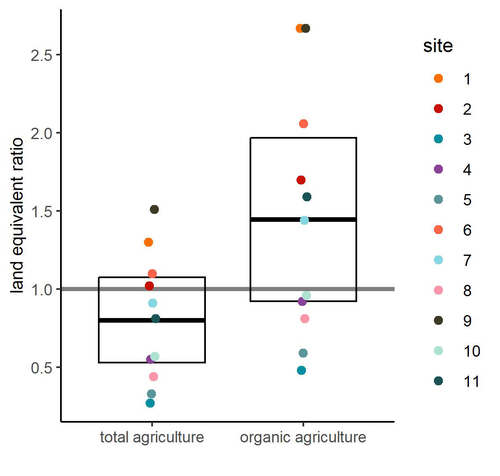
Crop productivity of Central European Permaculture is within the range of organic and conventional agriculture.Permaculture, a promising alternative to conventional agricultureRecommended by Aleksandra Walczyńska based on reviews by Julia Astegiano, Paulina Kramarz, Leda Lorenzo Montero and 1 anonymous reviewerAs mankind develops increasingly efficient and productive methods of agriculture and food production, we have reached a point where intensive agriculture threatens several aspects of life on Earth, negatively affecting biodiversity, carbon, nitrogen and phosphorus cycles and water reservoirs, while producing considerable amounts of greenhouse gases (Krebs and Bach, 2018). There was a need to develop farming methods that were friendly to both nature and people, producing good quality, healthy food without destroying the environment. The idea of permaculture, a concept of sustainable agriculture based on methods learned directly from nature, originated in the 1960s, invented and developed by Bruce Charles Mollison and David Holmgren (Mollison and Holmgren 1979, Mollison et al. 1991, Holmgren 2002). Although the idea of permaculture has attracted scientific interest, the representation in published studies is unbalanced in favour of positive ecological and sociological effects, with much less presence of rigorous experimental testing (Ferguson and Lovell 2014, Reiff et al. 2024a). Reiff et al. (2024b) provided the first large-scale empirical evidence of permaculture production outcomes for Central Europe. Based on results from 11 commercial permaculture sites, situated mostly in Germany but also in Switzerland and Luxembourg, the authors found that food production from permaculture sites was on average comparable to that from conventional and organic agriculture. The authors were very thorough in pointing out the issues that could potentially affect their results and which need further testing. Among these, the authors highlight the considerable variability between the 11 sites studied, which may suggest that different permacultures should differ in details according to their specificity - an interesting issue that definitely requires further study. The other factor that the authors point out that could have influenced the results and led to an underestimation of the real potential is the age of the permaculture sites. The sites from the study were relatively young, and their potential can be expected to increase with time. It is important to note that the results are mostly applicable to vegetables, as vegetable production accounted for 94% of production in the permaculture sites (followed by tree crops, 6%, and soft fruit production, 0.5%). There is therefore a need to include other types of crops produced in further studies of this type. To date, the results informing permaculture food production are urgently needed and should cover the potentially wide range of geographical regions and crops produced. The results of Reiff et al. (2025) show that rigorous testing of this issue is demanding, but the authors provide a very sound "road map" of further steps.
Literature: Ferguson R. S. and Lovell S. T. 2014. Permaculture for agroecology: design, movement, practice, and worldview. A review. Agronomy for Sustainable Development 34, 251-274. https://doi.org/10.1007/s13593-013-0181-6 Holmgren D. 2002. Permaculture: Principles & Pathways Beyond Sustainability. Holmgren Design Services, pp. 320. Krebs J. and Bach S. 2018. Permaculture – scientific evidence of principles for the agroecological design of farming systems. Sustainability 10, 3218, https://doi.org/10.3390/su10093218 Mollison B. C. and Holmgren D. 1979. Permaculture One: A Perennial Agricultural System for Human Settlements. Tagari Publications, pp. 136. Mollison B. C., Slay, R. M. and Jeeves A. 1991. Introduction to permaculture. Tagari Publications, pp. 198. Reiff J., Jungkunst H. F., Mauser K. M., Kampel S., Regending S., Rösch V., Zaller J. G. and Entling M. H. 2024a. Permaculture enhances carbon stocks, soil quality and biodiversity in Central Europe. Communications Earth & Environment 5, 305. https://doi.org/10.1038/s43247-024-01405-8 Reiff J., Jungkunst H. F., Antes N. and Entling M. H. 2024b. Crop productivity of Central European Permaculture is within the range of organic and conventional agriculture. bioRxiv, ver.2 peer-reviewed and recommended by PCI Ecology. https://doi.org/10.1101/2024.09.09.611985
| Crop productivity of Central European Permaculture is within the range of organic and conventional agriculture. | Julius Reiff, Hermann F. Jungkunst, Nicole Antes, Martin H. Entling | <p>Permaculture is a promising framework to design and manage sustainable food production systems based on mimicking ecosystems. However, there is still a lack of scientific evidence especially on the crop productivity of permaculture systems. In ... | | Agroecology | Aleksandra Walczyńska | 2024-09-09 13:37:04 | ||
21 Jan 2025
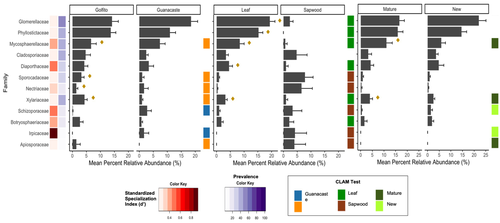
Exploring Rubiaceae fungal endophytes across contrasting tropical forests, tree tissues, and developmental stagesThe hidden diversity of fungal endophytes, associated with Rubiaceae of Costa Rican old-growth forestsRecommended by Melanie Roy based on reviews by Marion Boisseaux and 1 anonymous reviewerEndophytic fungi are expected to be hyperdiverse in tropical forests, and here is an article exploring their diversity, hidden in Rubiaceae leaves, in two old-growth forests of Costa Rica. Humberto Castillo-González et al. not only described their diversity, but also test for the impact of leaf development stage, tissue origin, and site location. They distinguish the different fungal lineages and do identify distinct indicators, showing that specialization of endophytic fungi could be related to other factors in tropical forests. This article is a great example of fungal ecology in the tropics, interacting at fine and large scale with a diversity of hosts. It also invites to discuss the high specialization observed in the tropics, and the ecology of old-growth forests in Costa Rica. References Humberto Castillo-González, Jason C. Slot, Stephanie Yarwood, Priscila Chaverri (2025) Exploring Rubiaceae fungal endophytes across contrasting tropical forests, tree tissues, and developmental stages. bioRxiv, ver.3 peer-reviewed and recommended by PCI Ecology https://doi.org/10.1101/2024.02.13.580172 | Exploring Rubiaceae fungal endophytes across contrasting tropical forests, tree tissues, and developmental stages | Humberto Castillo-González, Jason C. Slot, Stephanie Yarwood, Priscila Chaverri | <p>Fungal endophytes play a pivotal role in tropical forest dynamics, influencing plant fitness through growth stimulation, disease suppression, stress tolerance, and nutrient mobilization. This study investigates the effects of region, leaf devel... | | Agroecology, Biodiversity, Community ecology, Microbial ecology & microbiology, Mycology, Symbiosis | Melanie Roy | 2024-02-15 22:42:10 | ||
20 Jan 2025
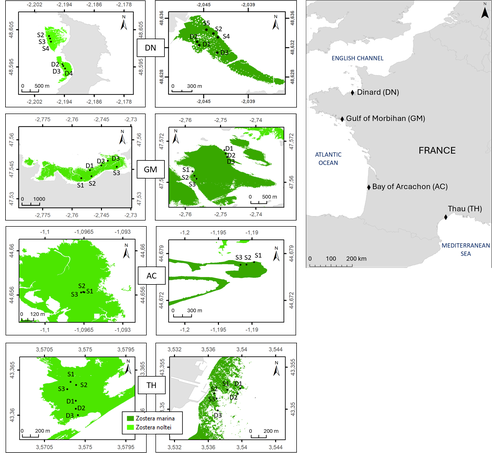
Regional and local variability in the morphometric traits of two emblematic seagrass species (Zostera marina and Zostera noltei) along the French coastImportance of Scale Considerations in Understanding Seagrass DynamicsRecommended by Antoine Vernay based on reviews by Gudrun Bornette and 2 anonymous reviewers
Seagrasses, particularly Zostera species, have been the subject of numerous studies due to their widespread distribution across the globe (Short et al., 2007), yet they have been in decline for several years as a result of global environmental changes (Touchette, 2007). While case studies and regional studies have been conducted, there remains a lack of information on how to scale these findings, particularly because of the heterogeneous nature of these habitats. In their study, Lacoste et al. examine the ecosystem trajectories of two Zostera species along a regional gradient spanning sites in the English Channel, the Atlantic Ocean, and the Mediterranean Sea. Their research is based on a recently published database, which offers valuable insights for comparing with other studies and serves as a resource for addressing future questions (Lacoste et al., 2024). This underscores the need for a global database to facilitate the integration of functional responses across studies, thus advancing our understanding of Zostera ecology on a larger scale. The multi-trait approach employed in their study provides a comprehensive view of population dynamics over a 1.5-year period, covering different seasons. Such studies highlight the complex responses of Zostera populations when considering environmental, seasonal, and geographical heterogeneity. Understanding these dynamics raises important questions about modeling, particularly in relation to the development of a more global database as previously mentioned. However, the review process has pointed out that the environmental data should be further refined to more rigorously support the presented results. Some statistical analyses could also benefit from improvements to ensure clearer and more explicit conclusions. These concerns are related to the challenges of sampling, the time required for such efforts, and the need to account for spatiotemporal variability. This study could serve as a foundational step for advancing our understanding of Zostera population dynamics on a global scale. In my opinion, despite the large ongoing scientific efforts, upscaling remains one of the major challenges for functional ecologists (Wood et al., 2024), particularly when plant habitats exhibit the kind of heterogeneity seen in Zostera, as demonstrated by Lacoste et al. in their work. References Élise Lacoste, Aurélien Boyé, Aline Blanchet-Aurigny, Nicolas Desroy, Isabelle Auby, Touria Bajjouk, Constance Bourdier, Nicolas Cimiterra, Céline Cordier, Amélia Curd, Lauriane Derrien, Élodie Foucault, Jean-Dominique Gaffet, Florian Ganthy, Loic Rigouin, Claire Rollet, Laura Soissons, Aurélien Tancray, Vincent Ouisse (2024) Regional and local variability in the morphometric traits of two emblematic seagrass species (Zostera marina and Zostera noltei) along the French coast. Zenodo, ver.3 peer-reviewed and recommended by PCI Ecology https://doi.org/10.5281/zenodo.10427767 Lacoste, E., Ouisse, V., Nicolas, D., Allano, L., Auby, I., Bajjouk, T., Bourdier, C., Caisey, X., de Casamajor, M.-N., Cimiterra, N., Cordier, C., Curd, A., Derrien, L., Droual, G., Dubois, F. S., Foucault, E., Foveau, A., Gaffet, J.-D., Ganthy, F., … Blanchet-Aurigny, A. (2024). A dataset of Zostera marina and Zostera noltei structure and functioning in four sites along the French coast over a period of 18 months. https://doi.org/10.5281/zenodo.14174128 Short, F., Carruthers, T., Dennison, W., & Waycott, M. (2007). Global seagrass distribution and diversity : A bioregional model. Journal of Experimental Marine Biology and Ecology, 350(1), 3‑20. https://doi.org/10.1016/j.jembe.2007.06.012 Touchette, B. W. (2007). The biology and ecology of seagrasses. Journal of Experimental Marine Biology and Ecology, 350(1), 1‑2. https://doi.org/10.1016/j.jembe.2007.06.013 Wood, G. V., Filbee-Dexter, K., Coleman, M. A., Valckenaere, J., Aguirre, J. D., Bentley, P. M., Carnell, P., Dawkins, P. D., Dykman, L. N., Earp, H. S., Ennis, L. B., Francis, P., Franco, J. N., Hayford, H., Lamb, J. B., Ling, S. D., Layton, C., Lis, E., Masters, B., … Wernberg, T. (2024). Upscaling marine forest restoration : Challenges, solutions and recommendations from the Green Gravel Action Group. Frontiers in Marine Science, 11. https://doi.org/10.3389/fmars.2024.1364263 | Regional and local variability in the morphometric traits of two emblematic seagrass species (*Zostera marina* and *Zostera noltei*) along the French coast | Élise Lacoste, Aurélien Boyé, Aline Blanchet-Aurigny, Nicolas Desroy, Isabelle Auby, Touria Bajjouk, Constance Bourdier, Nicolas Cimiterra, Céline Cordier, Amélia Curd, Lauriane Derrien, Élodie Foucault, Jean-Dominique Gaffet, Florian Ganthy, Loic... | <p><em>Z</em>Zostera marina and Zostera noltei are two foundation species that play a crucial role in the functioning of coastal ecosystems. They occur in a wide range of environmental conditions over a large geographical area in the northern hemi... | | Biogeography, Community ecology, Ecosystem functioning, Marine ecology, Morphometrics, Population ecology | Antoine Vernay | 2023-12-23 15:13:57 | ||
14 Jan 2025

Delayed dichromatism in waterfowl as a convenient tool for assessing vital ratesA cost-effective and non-invasive approach to estimating population dynamics in waterfowlRecommended by Huihuang Chen based on reviews by 2 anonymous reviewers
This article highlights a novel non-invasive method based on the "apparent sex ratios" that exploits delayed sexual importance in waterfowl populations. Unlike traditional capture-mark-recapture (CMR) technique, which is costly, invasive, and may disturb the target species, this method infers key population dynamics, such as adult survival rate and recruitment rate, by monitoring sex ratios in counts conducted during winter. Juvenile males that resemble adult females before molting provide a unique opportunity to estimate these vital rates. This method is cost-effective, minimizes disturbance to the species, and is particularly suitable for studying protected or invasive species. References Adrien Tableau, Iain Henderson, Sébastien Reeber, Matthieu Guillemain, Jean-François Maillard, Alain Caizergues (2024) Delayed dichromatism in waterfowl as a convenient tool for assessing vital rates. bioRxiv, ver.3 peer-reviewed and recommended by PCI Ecology https://doi.org/10.1101/2024.06.04.597326 | Delayed dichromatism in waterfowl as a convenient tool for assessing vital rates | Adrien Tableau, Iain Henderson, Sébastien Reeber, Matthieu Guillemain, Jean-François Maillard, Alain Caizergues | <p>Monitoring the number of individuals is by far the most popular strategy for studying the environmental factors that determine population dynamics and for measuring the effectiveness of management actions aimed at population recovery, control o... | | Biological control, Conservation biology, Demography, Life history, Population ecology, Statistical ecology | Huihuang Chen | 2024-06-07 17:39:34 |




