
Recommendation
 based on reviews by 2 anonymous reviewers
based on reviews by 2 anonymous reviewers
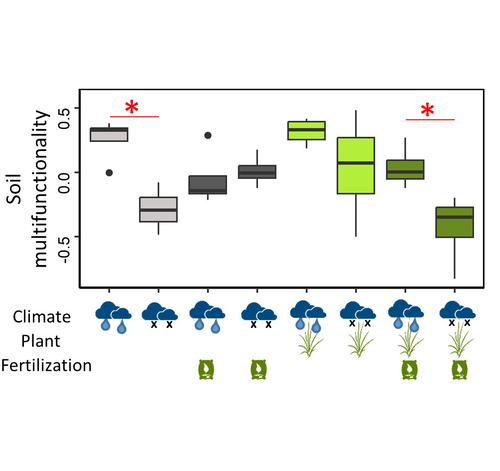
The ingredients of this study are classic in soil ecology and in the study of belowground-aboveground interactions: the presence of plants, draught and mineral fertilization (for the experimental treatments) and microbial carbon, microbial nitrogen, microbial phosphorus, substrate-induced respiration, cumulative extracellular enzyme activity, nitrogen mineralization potential, nitrification potential, denitrification potential (as a result of the treatments). It is interesting and useful to have tested all the combinations of the three treatments and the height variables (also in the form of a soil multifunctionality index) in the same study and to have been able to express hypotheses on the underlying mechanisms of interaction.
A key result is that mineral fertilization can reduce the soil ability to withstand draughts in terms of soil multifunctionality. This effect would be due to the increase in plant growth associated with fertilization, which reduces the availability of soil resources. This subsequently affects microbial diversity and soil multifunctionality. This confirms that the interactions between plants and soil microorganisms are complex and relevant for understanding and predicting the impact of climate and fertilization on soil functioning and the sustainability of plant-soil systems.
Although the study is rather fundamental, it has been designed to be relevant to grassland management and points to very general mechanisms that are likely to be relevant to arable land management. It would therefore be useful to repeat this work for interactions between a crop and its soil. Finally, it would be crucial to test the impact of heavy fertilization in intensive cropping systems on the resistance and resilience of soil functions to climate variability and climate changes.
A slightly disturbing fact is that the underlying interactions are probably so complicated that it seems so far impossible to me to make predictions about the ranking of the height combinations of treatments on each soil variable. But this complexity is clearly inherent to ecology and, in particular, plant-soil interactions.
References
Gabin Piton, Arnaud Foulquier, Lionel Bernard, Aurelie Bonin, Thomas Pommier, Sandra Lavorel, Roberto Geremia, Jean Christophe Clement (2025) Mineral fertilization reduces the drought resistance of soil multifunctionality in a mountain grassland system through plant-soil interactions. bioRxiv, ver.2 peer-reviewed and recommended by PCI Ecology https://doi.org/10.1101/2024.09.19.613911
DOI or URL of the preprint: https://doi.org/10.1101/2024.09.19.613911
Version of the preprint: 1
Find our reply to recommender and reviewers below after the ">".
Revision round #1
Decision for round #1 : Revision needed
Dear authors
I concur with the two reviewers to think that this manuscript is interesting and potentially useful. The reviewers have made many important comments that should be addressed to improve the manuscript quality. In particular, I think it would be useful for readers to see a better framing of the comparison between the plant and no-plant treatments and the addition of large amounts of nitrogen. In the discussion, this could lead to more reflection on the real ecological systems to which the experimental results could be compared and subsequently applied. A more minor point is that the whole experimental design and tested hypotheses depend on the test of interactions between treatments. This should be made clearer in the Results section. For example, the interactions tested in Table S2 should be described in the main text (or the absence of significant interaction).
Sebastien Barot
> We fully agree with your recommendation. Our experimental treatments (level of fertilization and choice of the grass species and the soil) aimed at representing managed grassland so we updated the text throughout the paper to clarify this point (eg.L11-13, L24-26, L54-59, L91-94, L95-96, L116-125, L395-396). Concerning the plant/no-plant treatment, its aim was to disentangle the potential role of plant in any interaction between fertilization and drought. We clarified this (eg. L96-99) and developed the introduction and discussion section on the plant role in the effect of fertilization and drought on the plant-soil system (L85-94, L370-384).
> Indeed, the test of our hypothesis relies on the statistical interaction between treatments in our model. Our text did describe the pattern emerging from these interactions when they were significant but we forgot to mention explicitly the significance of the interaction term in the text. We have now added this information directly in the main text (L252-254, L257-259, L306-307). Thanks for the comment.
Reviewer 1
Comments to the authors
This manuscript investigates the combined effects of drought and mineral nutrient addition (Nitrogen and Phosphorus) on soil multifunctionality in a mesocosm experiment with or without plants. The study addresses an important question given the increase in the frequency and severity of droughts, and investigates the response of variables both related to the soil functioning (among which, several enzyme activities related to C, N and P cycles, and microbial biomass C, N and P) and to the soil microbial diversity (both bacterial and fungal diversity, using DNA metabarcoding). Also, the authors use soil multifunctionality as an integrative variable of the response to the interactive effects of drought, nutrient addition and plants, which is very relevant in this context. The manuscript is well written, well structured, and well referenced. The methods are well described and the statistical analyses are appropriate.
Please see below some general and then some specific comments.
First, and this is my main comment, I think the authors should rephrase some parts of their manuscript to put their study less in the context of multiple and interacting global changes (see in particular L10-11, L23-24, L33-34, L81-82, L307-308, L369-370) and more in the context of the combined impact of drought and nutrient addition. I think this would be more appropriate and more adapted to the study conducted. Indeed, the level of nutrient added is closer to what is added in intensive sown grassland as it is clearly indicated by the authors (+150 kg N ha-1 and +80 kg P ha-1, see L122-124) than to the current and future atmospheric N deposition rates (e.g., see the papers from Galloway et al.).
>We agree with the reviewer comment that it sounds more appropriate to put our study in the specific context of managed grassland and their response to drought and fertilization rather than the general context of interacting global changes. We reword the text accordingly (L10-13,). We only keep the emphasis our contribution to the general issue of interacting global changes in the abstract opening (L23-24) and conclusion (L384-386).
Second, I think it could be useful if the authors could further explain, and at an early stage of the manuscript, what they exactly attempt to simulate with their “no plants” treatment, i.e. how much this is done in order to assess and distangle the effects mediated by plants and/or to mimick periods with no plants that could occur in agroecosystems (if the study is putted in a cropland context, as done at the very end of the manuscript, see L374-376, though here the system with Lolium perenne is closer to a managed grassland).
>The aim of the plant/no plant treatment was to disentangle the effect mediated by plants. We clarified this point at the end of the introduction (L96-99).
> As mentioned by the reviewer, the choice of the soil (from grazed mountain grassland) and of the plant (Lolium perenne, a very common grass in managed grassland) were done to represent conditions of managed grasslands where plant cover are never absent, or only for a short period of time for sown grasslands. Consequently, we agree that the discussion in the context of cropland at the end of the manuscript as mentioned by the reviewer was indeed out of scope. Therefore, we removed this sentence and rather stressed the implications of our results in the planted treatment that better represented grassland (L. 395-396).
Third, the authors have chosen to stop the watering during 32 days to simulate the drought event, and have collected the plant-soil samples at day 97, they could add a part in the discussion related to the duration of the drought/the experiment, and discuss the response of soil multifunctionality in relation to the severity of the drought event that they have simulated. Moreover, they have assessed their treatment effects on the resistance to drought, and, though the resilience is not directly assessed here, adding a short discussion on the duration of the effects and on the resilience of the soil multifunctionality based on results from the literature might be interesting.
> Reading the reviewer comment, we felt that the timing of our experiment was not clear enough. Our experiment start with 65 days to let plant grow, then we stopped watering during 32 days and collected the plant-soil samples at the end of this drought simulation. This correspond to the day 97 of the experiment (65+32), not 97 days after the start of the drought as it seems that the reviewer understood. We reword the experiment timing description to make it more clear (L127-129).
> We also added a section on the consequences of drought duration and resilience to further discuss our results (L325-335)
At the end of their introduction, the authors have decided to include their hypothesis in a Table (see Table 1), rather than in the main text, I personally think that it would be easier to read/follow if the hypotheses were included directly at the end of the introduction (but it might be a personal point of view).
> We followed the reviewer suggestion and developed the hypothesis in the main text (L99-101), and we removed the table.
In the methods, the authors indicate that “subsamples for enzymatic analyses were frozen at -20°C until analysis” (see L137-138), I guess this is not the case as enzymatic activities are classically assessed on fresh soils (i.e., on soil stored few days at 4°C prior to analysis). Could the authors please clarify this?
> The perfect practice would be indeed to measure enzyme activities on fresh soil within 24hrs after sampling (Burns et al. 2013). However, for experiments with many samples and variables, as this one, it is sometime not possible, and the samples must be stored. In our experiment, we preferred investing our time to measure microbial biomass on fresh soil within the 24hrs following sampling than focusing on enzymes, which are less sensitive. Therefore, soil samples were frozen at -20°C before being characterized with enzyme assays. While not ideal, freezing soil before enzyme assays has become a very common practice in soil biochemistry studies (eg. Baker and Allison (2017) and Dove et al. (2020)). Several studies assessed the consequences of different storage practices on enzyme activities (4°C, -20°C, -80°C and air drying, reviewed in in (Burns et al. 2013, Lane et al. 2022)). While they converged in showing that air-drying seems not appropriate, the best long-term storage among 4°, -20°C or -80C seems soil dependent. All long term storage seems to potentially influence the enzyme activities but the ability to discriminate different treatments seems relatively well conserved (Lane et al. 2022). As our study aimed to capture such relative difference rather than obtaining an absolute values of enzyme activity, as these values are any way standardized in our multifunctionality index, we thus argue that the use of frozen soil to measure enzyme activity in our study did not challenge our conclusion.
>All references are listed at the end of the response.
The authors could specify the way P was measured in their extracts (see L148-149).
> The method of Murphy and Riley (1962) was used. The information was added in the text.
For the DNA metabarcoding, please homogenize the information provided Table S1 and L204 (Table S1: v5-v6 regions, 16s rRNA gene; L204: v4 region of the 16s rRNA gene).
> Sorry. It was the v4 region, we homogenized the information.
The authors indicate “Model residuals were tested for normality and homoscedasticity and transformed when necessary”, I guess what they meant is that the data were transformed when necessary, please reword to clarify, and indicate which variables were transformed/which transformations were used.
> Indeed, our wording was incorrect. We reworded the sentence as “Model residuals were tested for normality and homoscedasticity and variables were log-transformed when necessary”. We also indicated the transformed variable in Table S2.
The authors indicate that the acronym used for Potential N mineralization is PNM (see L177), but then used the acronym NMP (see L197 and Table S2), so please homogenize.
> corrected
If the authors decide to keep their Table 1 with their hypothesis rather than including those in the main text, then the table on the effects of drought, fertilization and plant on bacterial and fungal community composition should be Table 2 and not Table 1 as currently indicated.
L368, the authors could omit the “1” before the title of the conclusion.
> done
The last figure of the Supplementary Material should be Figure S7 (and not SI Figure 5).
> corrected
The Table S2 is an important table of this manuscript, the authors should explain in the corresponding legend all the abbreviations used (most of the acronyms are easy to understand, or are already explained in the main text, but this is not the case for all, and it would help).
> We added the description of the abbreviations in the caption.
Reviewer 2
This work addressed the response of soil functioning and microbial composition in planted and unplanted mesocosms subjected to drought and fertilization. Soil function was characterized using soil multifunctionality, based on potential enzymatic activities as well as microbial biomass. The structure-function approach proposed provides strong data to test the different hypotheses, further reinforced by stoichiometric measurements. The question is timely, and has attracted many research efforts over the last decades.
I am in full favor of including plants in this type of study that has too often lacked them. However, showing that plant presence indeed has an effect on soil functioning feels somewhat trivial. This appears particularly clearly in the conclusion: there are few soils that are devoid of plants, and making a big deal out of plant presence vs absence seems to miss the point. I think that there is enough interesting data in this study to radically tone down focusing on the plant absence/presence treatment, and instead explore more in depth the mechanisms in the presence of plants.
This, in turn, requires to clarify the conceptual framework. The introduction should establish more clearly the angle chosen to address clarify plant-soil interactions in relation to environmental conditions. Perhaps this could help unfocus from plant presence/absence and refocus on less trivial aspects to punch up the discussion. Specifically: the authors first state soil microbes control most soil functions (L33), then that plants play a central role (L36), then emphasize the influence of soil abiotic factors on microbes (L37), without mentioning the influence of these abiotic controls on plants and on the plant-microbe interaction. Unfortunately, the latter is at the heart of what the discussion drives up to (L355-367). Table 1 reflects this: it explicits the conceptual framework, which is commendable, but still requires more work to be clear and reader-friendly. In addition, some logical articulations need to be made clearer, eg L45-54 (including syntax issues).
> We agree with the reviewer that the effect of plant presence on soil functioning response to global change is somewhat trivial and should not be the main message of the paper. We followed the reviewer recommendation and rather emphasized the results in the presence of plant, a condition that also better represents the study system (ie. grassland, see response to the other reviewer’s comment). We also worked on the introduction and discussion to clarified the framework and especially developed the importance of plant-soil interactions. Introduction and discussion were clarified and material was added as recommended by the reviewer, especially to address the response of plant to abiotic factors modified by fertilization and drought and their effect on soil functioning (L85-94, L370-384).
The paragraph about life history strategy addresses the nutrient availability niche dimension only (eg Fierer et al 2007 Ecology), however other dimensions could be considered, eg a water availability dimension (eg Barnard et al 2013 ISME), which would expand niche space and make sense when discussing bacterial G+:G- ratio.
> Thanks for this comment. The link between the nutrient and water niches is central in our hypothesis. This point was likely not explicit enough in the previous version. Therefore we developed this point and its consequences for the interaction between fertilization and drought in L59-66.
What is the rationale for the timeline of the experiment (30, 45, 32 days)?
> 30 days was the time necessary to obtain 2-3 leaves plants, strong enough to get planted. At 45 days, grasses started the tillering before its exponential growth, a stage often targeted to initiate the fertilization in the field. At day 65 grasses had greatly grown and covered all the mesocosms so we started to stop watering in the drought treatment. Then the 32 days period without water was defined based on the water dynamic in the different treatments. As fertilization highly promoted grass growth in the control treatment - which was watered twice a week - it highly stimulated plant water uptake and started to dry down the watered treatment with plant (Figure 2A, x axis). As we did not want the watered treatment with plant to be as dry as the drought treatment without plant, we decided to sample. This also fit with a moderate spring drought simulation as commonly reported in climate and global change studies (Karlowsky et al. 2018, Brunner et al. 2022, Möhl et al. 2023). We added all this information in the experimental design description (L120-131)
L159 microbial biomass elemental composition
> corrected
L163 The activity of 7 potential
> corrected
L170 methylumbelliferone
> corrected
L176 I doubt averaging 7 enzymes is enough to qualify for the name “total extracellular enzyme activity”, please rename.
> Yes we agree that these 7 enzymes do not cover the total enzyme activity in soil. We rename it to “cumulative enzyme activity”
L203 Change soil biodiversity to soil microbial diversity
> done
The first paragraph of the results omits any reference to any interaction. Why? These are the most interesting terms of the statistical model in a study that aims for drought x nutrient interaction. The first paragraph of the discussion should also much better explain the interaction that it points at.
> In the result section, our text did describe the pattern emerging from these interactions when they were significant but we forgot to mention explicitly the significance of the interaction term in the text. We have now added this information directly in the main text (L252-254, L257-259).
In the discussion section, we also revised the first paragraph where we highlighted our experimental interest in testing this interaction (L306-307.
The discussion surprisingly omits to mention rhizodeposition-related labile C flow from plants to soil microbes, the only occurrence I found of labile C being L348. No plants = no labile C for the microbes once the initial pool is gone = strong C limitation = little effect of fertilization on soil microbes, as shown in the results L277.
> The fact that plants added labile C into the soil and this way changed the stoichiometry of resource for microbe is indeed important. This was not emphasized enough in the previous version. We developed this point in the introduction (L86-87) and in the discussion (370-384) explaining the potential role of rhizodeposition in the limitation of microbes across our treatments.
The manuscript should be thoroughly checked for language (tenses, plurals, typos, syntax). L28 climate change mitigation, L39 stoichiometry, 43 However, some, 51 suggests, 53 SMF, 61 nutrients, L66 copiotrophs etc.
> Thank you very much for these corrections. We thoroughly checked the language before resubmitting.
> References for responses above
Baker, N. R., and S. D. Allison. 2017. Extracellular enzyme kinetics and thermodynamics along a climate gradient in southern California. Soil Biology and Biochemistry 114:82–92.
Brunner, M. I., A. F. Van Loon, and K. Stahl. 2022. Moderate and severe hydrological droughts in Europe differ in their hydrometeorological drivers. Water Resources Research 58:e2022WR032871.
Burns, R. G., J. L. DeForest, J. Marxsen, R. L. Sinsabaugh, M. E. Stromberger, M. D. Wallenstein, M. N. Weintraub, and A. Zoppini. 2013. Soil enzymes in a changing environment: current knowledge and future directions. Soil Biology and Biochemistry 58:216–234.
Dove, N. C., K. Arogyaswamy, S. A. Billings, J. K. Botthoff, C. J. Carey, C. Cisco, J. L. DeForest, D. Fairbanks, N. Fierer, R. E. Gallery, and others. 2020. Continental-scale patterns of extracellular enzyme activity in the subsoil: an overlooked reservoir of microbial activity. Environmental Research Letters.
Karlowsky, S., A. Augusti, J. Ingrisch, R. Hasibeder, M. Lange, S. Lavorel, M. Bahn, and G. Gleixner. 2018. Land use in mountain grasslands alters drought response and recovery of carbon allocation and plant-microbial interactions. Journal of Ecology 106:1230–1243.
Lane, J. M., C. S. Delavaux, L. Van Koppen, P. Lu, B. J. Cade-Menun, J. Tremblay, and L. D. Bainard. 2022. Soil sample storage conditions impact extracellular enzyme activity and bacterial amplicon diversity metrics in a semi-arid ecosystem. Soil Biology and Biochemistry 175:108858.
Möhl, P., M. Vorkauf, A. Kahmen, and E. Hiltbrunner. 2023. Recurrent summer drought affects biomass production and community composition independently of snowmelt manipulation in alpine grassland. Journal of Ecology 111:2357–2375.
Murphy, J., and J. P. Riley. 1962. A modified single solution method for the determination of phosphate in natural waters. Analytica chimica acta 27:31–36.
, posted 14 Dec 2024, validated 16 Dec 2024Dear authors
I concur with the two reviewers to think that this manuscript is interesting and potentially useful. The reviewers have made many important comments that should be addressed to improve the manuscript quality. In particular, I think it would be useful for readers to see a better framing of the comparison between the plant and no-plant treatments and the addition of large amounts of nitrogen. In the discussion, this could lead to more reflection on the real ecological systems to which the experimental results could be compared and subsequently applied. A more minor point is that the whole experimental design and tested hypotheses depend on the test of interactions between treatments. This should be made clearer in the Results section. For example, the interactions tested in Table S2 should be described in the main text (or the absence of significant interaction).
Sébastien Barot
Review for PCIEcology
Title
Plants reverse the positive effect of nutrient addition on the drought resistance of soil multifunctionality
Authors
Gabin Pinton, Arnaud Foulquier, Lionel Bernanrd, Aurélie Bonin, Thomas Pommier, Sandra Lavorel, Roberto Geremia, Jean-Christophe Clément
Comments to the authors
This manuscript investigates the combined effects of drought and mineral nutrient addition (Nitrogen and Phosphorus) on soil multifunctionality in a mesocosm experiment with or without plants. The study addresses an important question given the increase in the frequency and severity of droughts, and investigates the response of variables both related to the soil functioning (among which, several enzyme activities related to C, N and P cycles, and microbial biomass C, N and P) and to the soil microbial diversity (both bacterial and fungal diversity, using DNA metabarcoding). Also, the authors use soil multifunctionality as an integrative variable of the response to the interactive effects of drought, nutrient addition and plants, which is very relevant in this context. The manuscript is well written, well structured, and well referenced. The methods are well described and the statistical analyses are appropriate.
Please see below some general and then some specific comments.
First, and this is my main comment, I think the authors should rephrase some parts of their manuscript to put their study less in the context of multiple and interacting global changes (see in particular L10-11, L23-24, L33-34, L81-82, L307-308, L369-370) and more in the context of the combined impact of drought and nutrient addition. I think this would be more appropriate and more adapted to the study conducted. Indeed, the level of nutrient added is closer to what is added in intensive sown grassland as it is clearly indicated by the authors (+150 kg N ha-1 and +80 kg P ha-1, see L122-124) than to the current and future atmospheric N deposition rates (e.g., see the papers from Galloway et al.).
Second, I think it could be useful if the authors could further explain, and at an early stage of the manuscript, what they exactly attempt to simulate with their “no plants” treatment, i.e. how much this is done in order to assess and distangle the effects mediated by plants and/or to mimick periods with no plants that could occur in agroecosystems (if the study is putted in a cropland context, as done at the very end of the manuscript, see L374-376, though here the system with Lolium perenne is closer to a managed grassland).
Third, the authors have chosen to stop the watering during 32 days to simulate the drought event, and have collected the plant-soil samples at day 97, they could add a part in the discussion related to the duration of the drought/the experiment, and discuss the response of soil multifunctionality in relation to the severity of the drought event that they have simulated. Moreover, they have assessed their treatment effects on the resistance to drought, and, though the resilience is not directly assessed here, adding a short discussion on the duration of the effects and on the resilience of the soil multifunctionality based on results from the literature might be interesting.
At the end of their introduction, the authors have decided to include their hypothesis in a Table (see Table 1), rather than in the main text, I personally think that it would be easier to read/follow if the hypotheses were included directly at the end of the introduction (but it might be a personal point of view).
In the methods, the authors indicate that “subsamples for enzymatic analyses were frozen at -20°C until analysis” (see L137-138), I guess this is not the case as enzymatic activities are classically assessed on fresh soils (i.e., on soil stored few days at 4°C prior to analysis). Could the authors please clarify this?
The authors could specify the way P was measured in their extracts (see L148-149).
For the DNA metabarcoding, please homogenize the information provided Table S1 and L204 (Table S1: v5-v6 regions, 16s rRNA gene; L204: v4 region of the 16s rRNA gene).
The authors indicate “Model residuals were tested for normality and homoscedasticity and transformed when necessary”, I guess what they meant is that the data were transformed when necessary, please reword to clarify, and indicate which variables were transformed/which transformations were used.
The authors indicate that the acronym used for Potential N mineralization is PNM (see L177), but then used the acronym NMP (see L197 and Table S2), so please homogenize.
If the authors decide to keep their Table 1 with their hypothesis rather than including those in the main text, then the table on the effects of drought, fertilization and plant on bacterial and fungal community composition should be Table 2 and not Table 1 as currently indicated.
L368, the authors could omit the “1” before the title of the conclusion.
The last figure of the Supplementary Material should be Figure S7 (and not SI Figure 5).
The Table S2 is an important table of this manuscript, the authors should explain in the corresponding legend all the abbreviations used (most of the acronyms are easy to understand, or are already explained in the main text, but this is not the case for all, and it would help).
This work addressed the response of soil functioning and microbial composition in planted and unplanted mesocosms subjected to drought and fertilization. Soil function was characterized using soil multifunctionality, based on potential enzymatic activities as well as microbial biomass. The structure-function approach proposed provides strong data to test the different hypotheses, further reinforced by stoichiometric measurements. The question is timely, and has attracted many research efforts over the last decades.
I am in full favor of including plants in this type of study that has too often lacked them. However, showing that plant presence indeed has an effect on soil functioning feels somewhat trivial. This appears particularly clearly in the conclusion: there are few soils that are devoid of plants, and making a big deal out of plant presence vs absence seems to miss the point. I think that there is enough interesting data in this study to radically tone down focusing on the plant absence/presence treatment, and instead explore more in depth the mechanisms in the presence of plants.
This, in turn, requires to clarify the conceptual framework. The introduction should establish more clearly the angle chosen to address clarify plant-soil interactions in relation to environmental conditions. Perhaps this could help unfocus from plant presence/absence and refocus on less trivial aspects to punch up the discussion. Specifically: the authors first state soil microbes control most soil functions (L33), then that plants play a central role (L36), then emphasize the influence of soil abiotic factors on microbes (L37), without mentioning the influence of these abiotic controls on plants and on the plant-microbe interaction. Unfortunately, the latter is at the heart of what the discussion drives up to (L355-367). Table 1 reflects this: it explicits the conceptual framework, which is commendable, but still requires more work to be clear and reader-friendly. In addition, some logical articulations need to be made clearer, eg L45-54 (including syntax issues).
The paragraph about life history strategy addresses the nutrient availability niche dimension only (eg Fierer et al 2007 Ecology), however other dimensions could be considered, eg a water availability dimension (eg Barnard et al 2013 ISME), which would expand niche space and make sense when discussing bacterial G+:G- ratio.
What is the rationale for the timeline of the experiment (30, 45, 32 days)?
L159 microbial biomass elemental composition
L163 The activity of 7 potential
L170 methylumbelliferone
L176 I doubt averaging 7 enzymes is enough to qualify for the name “total extracellular enzyme activity”, please rename.
L203 Change soil biodiversity to soil microbial diversity
The first paragraph of the results omits any reference to any interaction. Why? These are the most interesting terms of the statistical model in a study that aims for drought x nutrient interaction. The first paragraph of the discussion should also much better explain the interaction that it points at.
The discussion surprisingly omits to mention rhizodeposition-related labile C flow from plants to soil microbes, the only occurrence I found of labile C being L348. No plants = no labile C for the microbes once the initial pool is gone = strong C limitation = little effect of fertilization on soil microbes, as shown in the results L277.
The manuscript should be thoroughly checked for language (tenses, plurals, typos, syntax). L28 climate change mitigation, L39 stoichiometry, 43 However, some, 51 suggests, 53 SMF, 61 nutrients, L66 copiotrophs etc.
